可見光與光譜#
人眼可見光的波長範圍極窄——約介於 380 至 750 奈米(nanometer,百萬分之一毫米)之間:
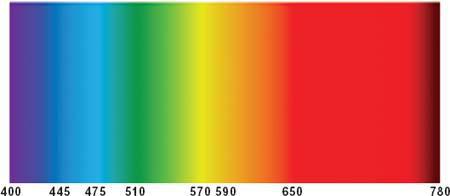
Figure 11: 可見光譜,標示各波長(單位:奈米)
| 波長範圍 | 對應感受 |
|---|---|
| < 380 nm | 紫外線(不可見) |
| 380–450 nm | 紫色 |
| 450–500 nm | 藍色 |
| 500–570 nm | 綠色 |
| 570–590 nm | 黃色 |
| 590–620 nm | 橙色 |
| 620–750 nm | 紅色 |
| > 750 nm | 紅外線(不可見) |
光線進入眼睛後,被視網膜(retina)後方的感光細胞吸收,轉成神經訊號送至大腦。色彩感受最終形成於大腦而非視網膜。
色彩感受其實是大腦的「錯覺」#
「同色異譜」現象(Metamerism)#
直覺上會以為「黃色」物體就是反射 580 nm 波長的光。事實上:
- 純單色(monochromatic)的 580 nm 黃光
- 與純紅光(620 nm)+ 純綠光(540 nm)等量疊加
兩者在我們眼中無法分辨——色彩感受完全相同。
電視螢幕正是利用這個原理——只用紅、綠、藍三種單色光的組合,就能模擬整個光譜。
物體的「黃」也並非單純黃光#
實際上「看起來黃色的物體」往往同時反射綠、紅、橙與黃光,並非只反射 580 nm 附近。
三色受體(Trichromacy)理論#
英國物理學家楊(Thomas Young)1801 年提出:色彩感的祕密不在光的物理性質,而在眼睛的解剖結構。後由馬克士威(James Clerk Maxwell, 1850s)與赫姆霍茲(Hermann von Helmholtz, 1860s)精煉,至今仍是視網膜運作的標準理論。
視網膜中的「錐細胞」(cones)#
色覺的基礎是三種光感色素分子,分別存在於三種錐細胞中:
| 錐細胞類型 | 峰值敏感度(波長) | 感受色域 |
|---|---|---|
| 短波(S) | ≈ 425 nm | 紫到藍,部分綠 |
| 中波(M) | ≈ 530 nm | 黃綠(中心),藍到橙 |
| 長波(L) | ≈ 565 nm | 黃綠到紅 |
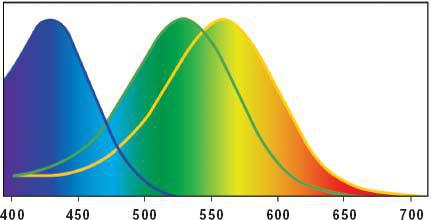
Figure 12: 三種錐細胞(短波、中波、長波)的標準化敏感度曲線(橫軸為波長 nm)
中波與長波錐細胞的峰值非常接近——這是人眼對黃綠區段特別敏感的原因。
錐細胞本身是「色盲的」#
每個錐細胞不知道自己吸收的光是什麼波長,它只記錄「總共吸收了多少光子」。例如:
- 短波錐細胞可能在「弱的紫光」與「強的綠光」中得到相同訊號
- 它無法分辨兩者
大腦透過比較三類錐細胞被激發的比例來推論色彩。
因此,許多不同的光譜分布只要產生相同的吸收比例,看起來就是相同顏色——這稱為「同色異譜」。
三維色彩空間#
我們的「色彩空間」是三維的(三類錐細胞的相對激發比例)——並非沿光譜線性分佈:
- 例如「粉紅色」並不對應任何單色光
- 而是來自紅光與藍光的特殊組合
- 因此存在於三維色彩空間中、卻不在彩虹光譜上
夜視的色盲#
當光線太弱,錐細胞無法運作。視網膜還有另一類細胞——桿細胞(rods):
- 對光極度敏感(單一光子即可激發)
- 峰值在 500 nm 左右(藍綠色)
- 只有一種類型,因此無法產生色彩感
對不同波長的不平均敏感度#
為何黃色看起來最亮?#
| 錐細胞 | 數量 |
|---|---|
| 短波(藍) | 最少 |
| 中波(綠) | > 短波的 10 倍 |
| 長波(紅黃) | 更多 |
因此眼睛對長波光(黃、紅)效率較高——同樣強度下,黃光看起來比藍光亮。 我們最大日視敏感度在 555 nm(黃綠色)。
但藍光的能量其實更高#
馬格努斯(Hugo Magnus)以為紅光能量最高,是徹底錯誤——波長越短,能量越高:
- 紅光:能量最低
- 黃 > 紅
- 綠、藍 > 黃
- 紫外線:能量極高,足以傷害皮膚
黃綠區段的精準辨色能力#
由於中波與長波錐細胞峰值極近,黃綠區的微小波長差異就會造成兩種錐細胞激發比例的明顯改變:
- 黃色區:可分辨僅差 1 nm 的色相
- 藍色區:辨識力不到上述的 1/3
- 紅色邊緣:比藍色更差
這種色彩空間的不對稱性,正是某些色彩劃分比其他劃分「更自然」的解剖學基礎。
色盲(Color Blindness)#
| 類型 | 定義 | 比例 |
|---|---|---|
| 紅綠色盲(dichromacy) | 缺中波或長波錐細胞 | 男性約 8%、女性約 0.45% |
| 藍黃色盲(tritanopia) | 缺短波錐細胞 | 約 0.002% |
| 全色盲(monochromat) | 只剩一種錐細胞 | 極罕見 |
| 桿細胞單色(rod monochromat) | 三種錐細胞皆無,只有桿細胞 | 更罕見 |
我們對色盲者實際看到什麼仍知之甚少——少數一眼正常一眼色盲者報告,色盲眼會看到「黃」與「藍」。但因神經連接可能也異常,這些報告不易精準解讀。
色覺的演化#
演化路徑#
人類色覺與舊大陸猴及類人猿同源,可追溯約 3,000–4,000 萬年。多數哺乳動物只有兩種錐細胞(雙色視覺):
| 演化階段 | 出現的錐細胞 | 時代 |
|---|---|---|
| 1(最早) | 短波(紫藍) | 哺乳動物前 |
| 2 | 中波(綠) | 哺乳動物階段 |
| 3 | 長波(黃綠 / 紅) | 約 3,000–4,000 萬年前 |
為何長波錐細胞會出現?#
一次基因複製事件,把原本的中波(綠)受體分裂為兩個相鄰受體——新的那個略偏黃。 這對「在綠葉背景下偵測黃色成熟果實」極為理想。
一位科學家戲言:「人類的三色視覺,不過是某些結果樹發明出來、用來繁殖自己的工具。」 這類熱帶樹的果實太大,鳥類無法搬運;牠們需要靠猴子吃下後吐出種子或在他處排便傳播。猴子之於彩色水果,正如蜜蜂之於花朵。
對馬格努斯理論的反駁#
從數百萬年的長尺度看,色覺演化的方向正好與馬格努斯所主張相反:
並非「從紅光感受先發展,向紫端推進」,而是「從藍紫感受開始,向紅端推進」。
大腦的「Photoshop」:色彩恆常性#
錐細胞只是色彩感受的起點——真正關鍵的是大腦的後處理。
為何便宜相機在室內燈光下色偏?#
並非相機在說謊:
- 黃色白熾燈下,物體確實反射更多黃光、藍光變灰
- 任何客觀儀器都會這麼測
但我們的大腦會把它「修正回來」——這個過程稱為色彩恆常性(color constancy)。
從演化角度看:若同一棵樹的水果中午與傍晚顏色不同,色彩就無法成為可靠的辨識線索——反而是阻礙。 大腦因此演化出「即時修正」機制,讓我們在不同光源下仍見到大致穩定的色彩。
比白平衡更精密#
數位相機的白平衡是機械式的;大腦的常態化則仰賴經驗、記憶與習慣。
Giessen 實驗(2006):香蕉的記憶#
德國 Giessen 大學實驗:
- 顯示螢幕上一片隨機色塊,請受試者調整顏色直到「完全灰色」
- 結果:受試者真的調到中性灰
- 同樣任務,但這次螢幕上是香蕉的圖
- 結果:受試者調到「略偏藍」才覺得香蕉看起來是灰
- 也就是說:當香蕉實際上已經是中性灰時,受試者仍覺得它略帶黃色
這證明:大腦會根據長期記憶與物體辨識主動「補正」色彩感受——它記得香蕉是黃的,所以把感受推往黃的方向。
語言介入色彩處理的可能機制#
語言對色彩感受的影響,極可能就發生在「常態化與補償」這個層次。
大腦在判斷「兩個顏色有多相似」時,仰賴儲存的記憶與已建立的範疇分類。 母語提供的色彩概念與差異訓練,正是這套記憶與分類的一部分。
雖然詳細機制尚未明朗,但這個附錄為前述「俄羅斯藍色」、「英語半腦不對稱」、「Mandarin MRI」等實驗結果,提供了神經解剖學上的合理基礎:
語言不是物理鏡片——它不會折射光線。 但它可能塑造大腦進行「色彩常態化」時所依賴的記憶結構—— 這正是語言能夠成為「感知鏡頭」的科學基礎。