病毒的基本結構與分類#
病毒(virus)是極微小(通常 20–30 nm)的感染性顆粒,無法在宿主細胞外自行複製。自由存在的病毒顆粒稱為病毒體(virion),由核酸(RNA 或 DNA)外包蛋白質外殼——衣殼(capsid)——所構成,兩者合稱核衣殼(nucleocapsid)。部分病毒另有脂蛋白套膜(envelope),上面嵌有抗原性糖蛋白。
病毒分為兩大類:
- DNA 病毒:痘病毒(天花)、疱疹病毒(水痘、帶狀疱疹、唇疱疹、腺熱)、腺病毒、乳突病毒(疣)
- RNA 病毒:正黏病毒(流感)、副黏病毒(麻疹、腮腺炎)、風疹病毒、狂犬病毒、小 RNA 病毒(普通感冒、脊髓灰質炎)、反轉錄病毒(retrovirus)(AIDS、T 細胞白血病)、砂粒病毒(Lassa fever)、肝 DNA 病毒(B 型肝炎)
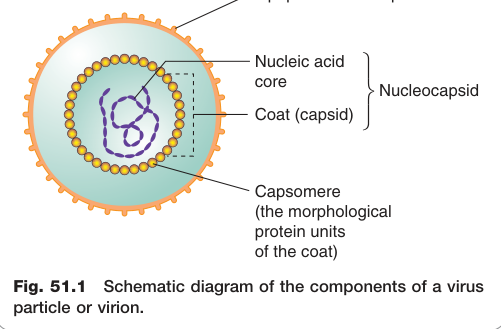
Figure 51.1:病毒顆粒(virion)的結構組成示意圖
病毒的生命週期#
病毒本身無代謝機制,必須入侵宿主細胞後才能複製。
進入宿主細胞#
病毒套膜或衣殼上的多肽結合位與宿主細胞膜上的受體(如細胞激素受體、神經傳遞物受體、離子通道等)結合,隨後多以受體媒介內吞方式進入細胞;進入後宿主酵素(多為溶酶體)去除病毒外殼,釋出核酸。
DNA 病毒的複製#
病毒 DNA 進入宿主細胞核,由宿主 RNA 聚合酶轉錄為 mRNA,再翻譯出病毒特異性蛋白(包括合成更多病毒 DNA 的酵素及結構蛋白),最後組裝完整病毒體並以出芽或細胞裂解方式釋出。
RNA 病毒的複製#
病毒攜帶的酵素從 RNA 模板合成 mRNA,由宿主核糖體翻譯出 RNA 聚合酶及結構蛋白,再組裝釋出;通常不涉及宿主細胞核(流感病毒為例外)。
反轉錄病毒的複製#
反轉錄病毒攜帶反轉錄酶(reverse transcriptase,RNA 依賴性 DNA 聚合酶),將病毒 RNA 反轉錄為 DNA。此 DNA 嵌入宿主基因組,成為前病毒(provirus),之後轉錄出新的病毒 RNA 及 mRNA,製造並釋出新的病毒體。許多反轉錄病毒可在不殺死宿主細胞的情況下複製,或長期以潛伏形式存在。
潛伏性感染(如唇疱疹、帶狀疱疹)的週期性復發,正是因為病毒前病毒 DNA 被嵌入宿主基因組後,在特定誘因下重新活化。
宿主對病毒的防禦#
先天與適應性免疫#
- 皮膚屏障:完整皮膚可阻擋多數病毒;破損皮膚和黏膜則較脆弱。
- T 細胞免疫:受病毒感染的細胞在表面呈現病毒胜肽與主要組織相容性複合體(MHC)第一類分子的複合物,由 **CD8+ 殺手 T 細胞(cytotoxic T lymphocyte, CTL)**辨識後殺滅——透過釋放溶胞蛋白(穿孔素、顆粒酶)或活化 Fas「死亡受體」誘發細胞凋亡。
- 自然殺手細胞(NK cell):當病毒下調 MHC 分子以逃避 CTL,NK 細胞即對缺乏正常 MHC 表現的細胞發動攻擊。
- 基因靜默:病毒複製產生的雙股 RNA 片段(siRNA)可靜默其對應基因,中斷病毒蛋白合成週期。
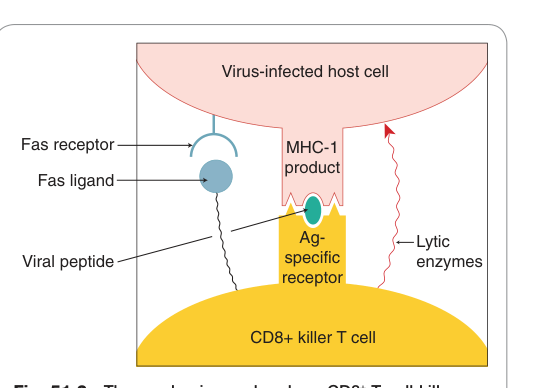
Figure 51.2:CD8+ 細胞毒殺 T 細胞(CTL)辨識 MHC-I/病毒胜肽複合物後,透過穿孔素、顆粒酶或 Fas 途徑殺死病毒感染宿主細胞的機制
病毒逃避宿主防禦的策略#
- 抑制細胞激素:部分痘病毒表達偽受體,競爭性結合 IL-1、TNF-α、干擾素(IFN),阻止其到達免疫細胞。
- 干擾 MHC 呈現:腺病毒、疱疹病毒、人類巨細胞病毒等抑制抗原胜肽的產生或 MHC–胜肽複合物的表現,使 CTL 無法偵測受感染細胞。
- 干擾細胞凋亡:腺病毒、巨細胞病毒、EB 病毒可破壞細胞凋亡途徑以確保自身存活。
- 偽裝 MHC:巨細胞病毒(cytomegalovirus)表達 MHC 第一類的同源物,欺騙 NK 細胞使其不攻擊。
大多數病毒感染在免疫功能正常的宿主中可自行緩解;免疫妥協(immunocompromised)宿主則難以控制病情。
HIV 與 AIDS#
**人類免疫缺乏病毒(human immunodeficiency virus, HIV)**是 RNA 反轉錄病毒。HIV-1 為全球 AIDS 主要病原,HIV-2 毒性較低且主要局限於非洲部分地區。
HIV 的入侵機制#
HIV 病毒體表面糖蛋白 gp120 與宿主細胞上的 CD4 糖蛋白結合;另一糖蛋白 gp41 則促使病毒套膜與細胞膜融合。輔助共受體為:
- CCR5:趨化因子受體,是多數 HIV 株入侵的主要共受體
- CXCR4:在晚期感染中越來越重要
帶有 CCR5 功能性缺失突變的個體對 HIV 感染具有較高抵抗力,這是 CCR5 拮抗劑開發的生物學基礎。
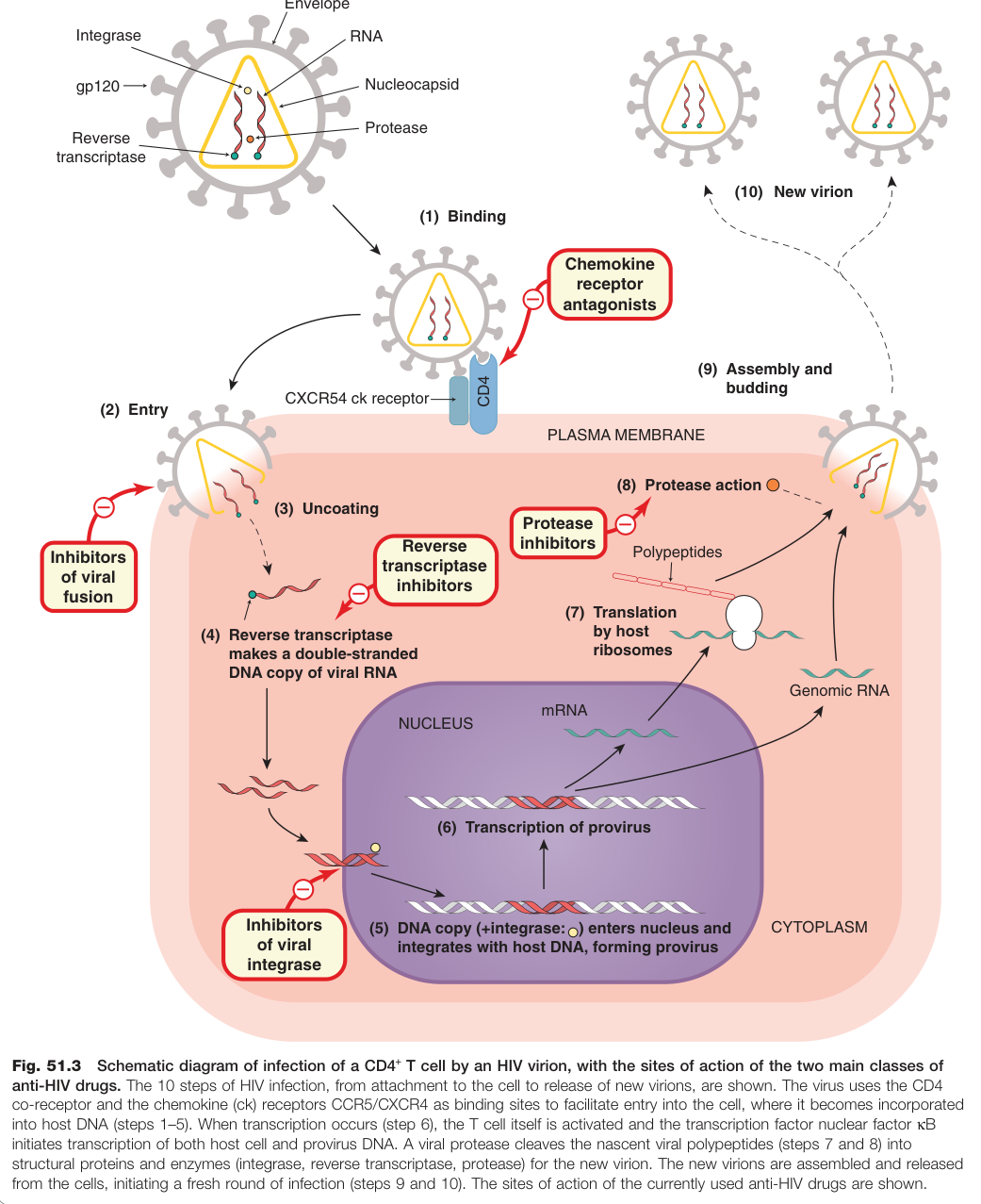
Figure 51.3:HIV 病毒體感染 CD4+ T 細胞的示意圖,標示兩大類抗 HIV 藥物的作用位點
HIV 感染的病程#
- 急性期:病毒血症(viraemia)上升、廣泛散播,引發類流感症狀;數週內因 CTL 反應使病毒量下降。
- 臨床潛伏期(中位數約 10 年):病毒在淋巴結靜默複製,CD4+ T 細胞數量持續下降。
- AIDS 期:免疫反應崩潰,出現伺機性感染(如肺囊蟲肺炎、結核病)、神經系統病變、惡性腫瘤等;未治療者通常在 2 年內死亡。
HIV 每日可產生多達 10¹⁰ 個新病毒顆粒,且突變率極高,使 CTL 及藥物均難以長期追蹤其靶點。
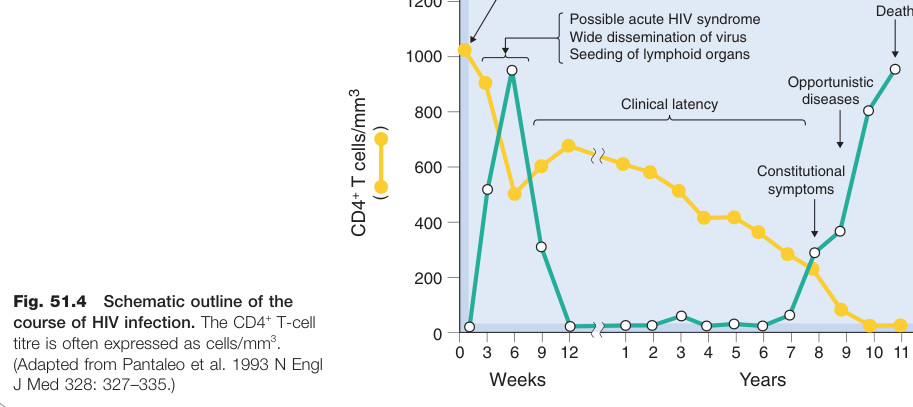
Figure 51.4:HIV 感染病程概況,顯示 CD4+ T 細胞計數與病毒量隨時間的變化
抗病毒藥物#
由於病毒劫持宿主的代謝機制,難以找到對病原具選擇性的藥物。現有抗病毒藥多針對病毒特有的酵素,且通常只在病毒複製期間有效。
反轉錄酶抑制劑#
核苷類似物(Nucleoside Reverse Transcriptase Inhibitors, NRTIs)#
代表藥物:齊多夫定(zidovudine, AZT)、阿巴卡韋(abacavir)、拉米夫定(lamivudine)、替諾福韋(tenofovir)等。
作用機制:
- 被宿主細胞酵素磷酸化為 5’-三磷酸衍生物
- 競爭病毒反轉錄酶,摻入生長中的病毒 DNA 鏈後導致鏈終止(chain termination)
- 對宿主 α-DNA 聚合酶影響較小,但對線粒體 γ-DNA 聚合酶有一定影響,是部分副作用的來源
齊多夫定為首個上市的抗 HIV 藥,可口服或靜脈注射,血漿半衰期 1 小時,胞內活性三磷酸形式半衰期 3 小時,CSF 濃度約為血漿的 65%。給予分娩母親及新生兒可降低母嬰傳播超過 20%。長期使用易產生抗藥性。
常見副作用(此類藥物共有):
- 胃腸道不適(噁心、嘔吐)
- 血液疾病(貧血、嗜中性白細胞減少)
- 中樞神經系統症狀(失眠、暈眩、頭痛)
- 乳酸中毒、脂肪代謝異常(lipodystrophy)
非核苷類反轉錄酶抑制劑(Non-Nucleoside RTIs, NNRTIs)#
代表藥物:奈維拉平(nevirapine)、依法韋侖(efavirenz)。
- 化學結構多樣,直接結合反轉錄酶催化位附近使其失活
- 多為 CYP450 酵素的誘導劑、受質或抑制劑,有藥物相互作用
- 奈維拉平有良好口服生物利用度,可預防 HIV 母嬰傳播
- 依法韋侖口服,半衰期約 50 小時(每日一次),主要副作用:失眠、噩夢、精神症狀;具致畸性,早孕期禁用
蛋白酶抑制劑(Protease Inhibitors, PIs)#
代表藥物:利托那韋(ritonavir)、沙奎那韋(saquinavir)、茚地那韋(indinavir)、達魯那韋(darunavir)、洛匹那韋(lopinavir)等。
HIV 前病毒轉錄的 mRNA 翻譯為兩條無活性多蛋白;病毒特有蛋白酶將多蛋白切割為結構及功能蛋白。蛋白酶抑制劑與切割位點結合,阻斷此步驟。
- 利托那韋常與其他蛋白酶抑制劑聯用以增強效果(藥物動力學增強)
- 口服,血漿半衰期 3–5 小時,主要由糞便排出
副作用:胃腸道不適、血液疾病、高血糖、脂肪代謝異常。
DNA 聚合酶抑制劑#
代表藥物:阿昔洛韋(aciclovir)、泛昔洛韋(famciclovir)、伐昔洛韋(valaciclovir,阿昔洛韋前藥)、更昔洛韋(ganciclovir)、西多福韋(cidofovir)、膦甲酸(foscarnet)。
阿昔洛韋作用機制:
- 被病毒胸腺嘧啶激酶(病毒酵素活性遠高於宿主酵素)優先磷酸化為單磷酸
- 宿主細胞激酶再轉換為三磷酸(活性形式)
- 抑制病毒 DNA 聚合酶,對宿主酵素效力低約 30 倍
- 摻入 DNA 鏈後導致鏈終止
主要適應症:
- 水痘帶狀疱疹感染(chickenpox, shingles)
- 單純疱疹感染(生殖器疱疹、疱疹腦炎)
- 免疫妥協患者的預防性用藥
副作用少見;靜脈輸注時需注意滲漏性炎症及腎功能障礙(緩慢輸注可降低風險)。
阿昔洛韋選擇性高的關鍵在於:其磷酸化主要由病毒胸腺嘧啶激酶完成,因此只在受感染細胞中才能被充分活化。
神經氨酸酶抑制劑與病毒脫殼抑制劑#
流感病毒(RNA 病毒)入侵需先附著宿主細胞的唾液酸(sialic acid);新複製的病毒體則透過**神經氨酸酶(neuraminidase)**切斷與宿主細胞的連結而釋出。
神經氨酸酶抑制劑:
- 扎那米韋(zanamivir):吸入粉劑,對甲型及乙型流感均有效
- 奧司他韋(oseltamivir):口服,對甲型及乙型流感均有效
兩者需在感染早期使用,副作用主要為胃腸道不適。
病毒脫殼抑制劑:
- 金剛烷胺(amantadine):阻斷流感病毒 M2 離子通道,抑制病毒脫殼(uncoating);僅對甲型流感有效,現已少用;副作用有暈眩、失眠(也輕微用於帕金森病)
其他作用機制的藥物#
| 藥物 | 機制 | 用途 |
|---|---|---|
| 恩夫韋肽(enfurvirtide) | 抑制 HIV 與宿主細胞融合 | HIV 組合療法(耐藥或不耐受時) |
| 拉替拉韋(raltegravir) | 抑制 HIV 整合酶(integrase),阻止前病毒嵌入宿主基因組 | HIV 耐藥病例的組合療法 |
| 馬拉維羅(maraviroc) | CCR5 趨化因子受體拮抗劑,阻斷 HIV 入胞 | 確認為 R5 株的 HIV 感染 |
馬拉維羅僅對使用 CCR5 入胞的 HIV 株有效,使用前需確認病毒株型別。
生物製劑與免疫調節劑#
免疫球蛋白(immunoglobulin):
- 集合免疫球蛋白含多種病毒抗體,可在症狀前使用以預防或減輕麻疹、風疹、肝炎、狂犬病等
- 高免疫球蛋白(hyperimmune globulin)針對特定病毒(B 型肝炎、水痘帶狀疱疹、狂犬病)
帕利珠單抗(palivizumab):抗呼吸道合胞病毒(RSV)糖蛋白的單株抗體,用於嬰兒預防性肌肉注射。
干擾素(interferon, IFN):
- 由哺乳動物細胞誘導合成,現多以重組 DNA 技術生產
- IFN-α、β 由 B/T 淋巴球、巨噬細胞、纖維芽細胞在病毒刺激下產生
- 與宿主細胞膜上的神經節苷脂受體結合,誘導酵素抑制病毒 mRNA 翻譯
- IFN-α-2a 用於 B 型肝炎及 AIDS 相關卡波西肉瘤;IFN-α-2b 用於 C 型肝炎
- **聚乙二醇化干擾素(pegylated IFN)**半衰期更長
- 副作用:發燒、倦怠、頭痛、肌痛(流感樣);長期使用可能造成骨髓抑制、皮疹、脫髮及心肝甲狀腺功能異常
三氮唑核苷(ribavirin/tribavirin):干擾病毒 mRNA 合成,主要以霧化或口服治療 RSV 感染及 C 型肝炎;對 Lassa 熱亦有效。
肌苷普蘭諾貝(inosine pranobex):干擾病毒核酸合成,兼具免疫增強作用,用於皮膚及黏膜疱疹感染。
HIV 的組合療法#
高效抗反轉錄病毒療法(HAART)#
單一藥物因 HIV 突變率極高而易產生抗藥性,組合療法已成標準。典型的 **HAART(highly active antiretroviral therapy)**方案:
- 兩種核苷 RTI + 一種非核苷 RTI,或
- 兩種核苷 RTI + 一至兩種蛋白酶抑制劑
- 必要時加入整合酶抑制劑、CCR5 拮抗劑或融合抑制劑
HAART 的原則:
- 監測血漿病毒量與 CD4+ 細胞計數
- 在免疫缺陷明顯前即開始治療
- 盡量將血漿病毒量降至不可測水平
- 若病毒量上升,立即更換方案
HAART 可顯著延長患者存活,但病毒並未被消滅——HIV 以潛伏形式藏匿於記憶 T 細胞的宿主基因組中,停藥即可能重新活化。長期治療所需的高度依從性、藥物相互作用及多重副作用均是挑戰。
新型抗病毒藥物的展望#
1990 年代初期僅有五種抗病毒藥;二十年後已增加近十倍。未來方向包括:
- CXCR4 抑制劑:類似馬拉維羅的 CCR5 拮抗策略
- siRNA 技術:靶向病毒特定基因進行靜默
- HIV 疫苗:抗原漂移(antigenic drift)使研發困難重重——超過 40 個候選疫苗已完成臨床試驗,至今仍無批准上市的 HIV 疫苗;研究方向轉向整合先天與適應性免疫的設計