概述#
化學治療(chemotherapy) 一詞最初由 Paul Ehrlich 在 20 世紀初提出,指以合成化學物質或抗生素殺滅或抑制病原體,同時對宿主毒性最小。近年來此詞涵蓋範圍已擴大至包括抗腫瘤藥物。
人類易感的病原體涵蓋病毒、細菌、原蟲、真菌及蠕蟲。選擇性毒性(selective toxicity) 的實現,有賴於找到病原體與宿主之間可供利用的生化差異。
「抗生素革命」始於 1940 年代盤尼西林問世,是近代醫學史上最重要的治療突破之一。然而,細菌正快速演化出耐藥機制,抗藥性問題已成為當代臨床的重大威脅。
化學治療的分子基礎#
病原體的分類與挑戰#
- 原核生物(prokaryotes):無核細胞,如細菌。
- 真核生物(eukaryotes):有核細胞,如原蟲、真菌、蠕蟲。
- 病毒:必須借用宿主細胞的代謝機制複製,是最難選擇性攻擊的對象。
- 癌細胞:與正常宿主細胞高度相似,選擇性毒性最難實現。
- 朊病毒(prions):目前無有效解毒劑。
細菌的基本結構#
細菌的生化反應依目標性質分為三類:
- Class I:利用葡萄糖等碳源生成 ATP 及簡單碳化合物(前驅物)。
- Class II:利用前驅物與能量合成胺基酸、核苷酸、磷脂、胺基糖等小分子。
- Class III:將小分子組裝為大分子——蛋白質、RNA、DNA、多醣及肽聚糖(peptidoglycan)。
其他潛在靶點包括:細胞膜、微管、食物液泡及肌肉纖維(後三者見於較高等病原體)。
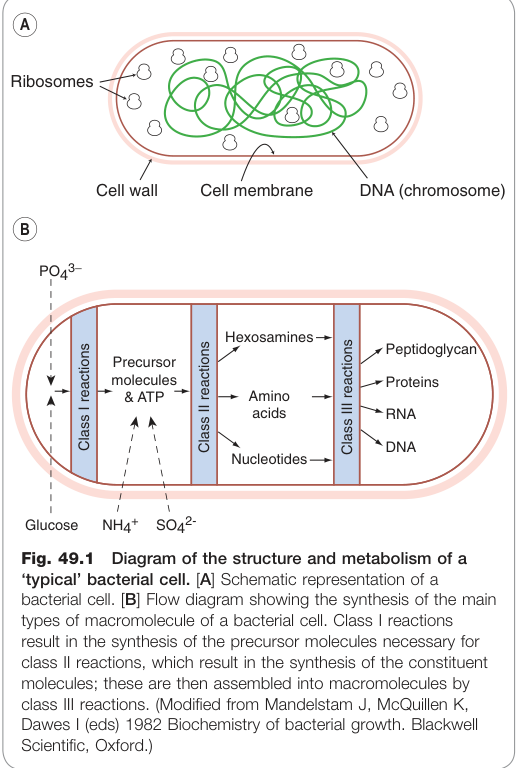
Figure 49.1:典型細菌細胞的結構示意圖及主要大分子(蛋白質、RNA、DNA、多醣、肽聚糖)的合成流程
各類反應作為化療靶點#
Class I 反應(不理想)#
- 細菌與人類細胞使用相同的能量代謝途徑(Embden–Meyerhof 途徑、TCA 循環)。
- 即使抑制葡萄糖氧化,細菌仍可利用胺基酸、乳酸等替代能源。
Class II 反應(較佳靶點)#
某些代謝途徑僅存在於細菌,不存在於人類。
葉酸(Folate)途徑#
- 人類無法自行合成葉酸,必須從食物攝取,且有主動運輸機制。
- 大多數細菌及瘧疾原蟲缺乏轉運機制,必須從頭合成(de novo)葉酸。
- 磺胺類藥物(sulfonamides):含磺胺基團,是 PABA 的結構類似物,競爭性抑制葉酸合成酶 → 抑菌(bacteriostatic),需宿主防禦配合才有效。
- 二氫葉酸還原酶(dihydrofolate reductase, DHFR) 抑制劑:同一酵素在不同生物體的敏感性差異極大(見下表):
| 抑制劑 | 人類 IC₅₀ (µmol/l) | 原蟲 IC₅₀ | 細菌 IC₅₀ |
|---|---|---|---|
| 甲氧苄啶(trimethoprim) | 260 | 0.07 | 0.005 |
| 乙胺嘧啶(pyrimethamine) | 0.7 | 0.0005 | 2.5 |
| 甲氨蝶呤(methotrexate) | 0.001 | ~0.1 | 無活性 |
嘧啶與嘌呤類似物#
- 氟尿嘧啶(fluorouracil):轉化為偽核苷酸,干擾胸苷酸合成,用於癌症治療。
- 氟胞嘧啶(flucytosine):在真菌細胞內去胺基化為氟尿嘧啶,人類細胞轉化量少,具選擇性。
Class III 反應(重要靶點)#
肽聚糖合成#
- 肽聚糖(peptidoglycan)僅存在於原核細胞的細胞壁,真核生物無此結構。
- 革蘭氏陰性菌:單層肽聚糖;革蘭氏陽性菌:多達 40 層。
- 抗生素在不同步驟阻斷合成:
| 抗生素 | 作用位置 |
|---|---|
| 環絲胺酸(cycloserine) | 競爭性抑制 D-丙胺酸加至 N-乙醯胞壁酸的初始三肽 |
| 萬古黴素(vancomycin) | 阻止構建模塊從脂質載體釋放 |
| 桿菌肽(bacitracin) | 干擾 C₅₅ 脂質載體的再生 |
| β-內醯胺類(β-lactams)(盤尼西林、頭孢素等) | 抑制最終轉肽反應,阻止交叉鍵形成 |
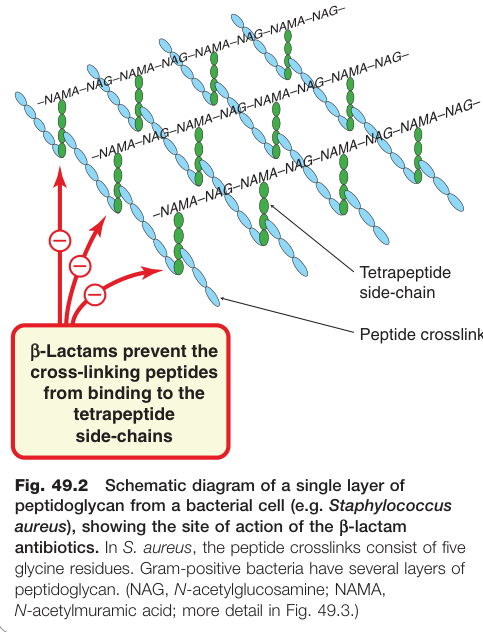
Figure 49.2:金黃色葡萄球菌肽聚糖單層結構示意圖,標示 β-內醯胺類抗生素的作用位置(轉肽交叉鍵形成步驟)
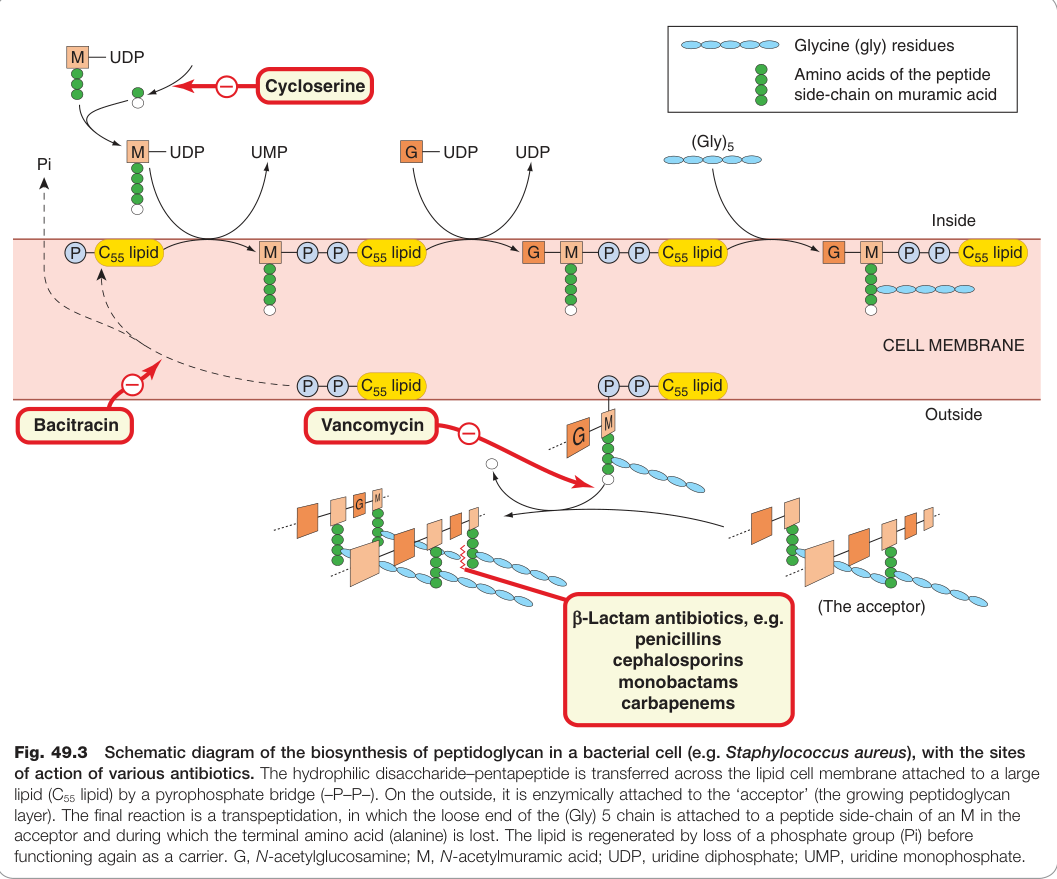
Figure 49.3:細菌肽聚糖生物合成流程(以金黃色葡萄球菌為例),標示環絲胺酸、萬古黴素、桿菌肽及 β-內醯胺類等抗生素的作用節點
蛋白質合成#
原核(70S = 50S + 30S)與真核(80S = 60S + 40S)核糖體結構不同,是抗生素選擇性的基礎。
抗生素作用於翻譯的各個步驟:
- 四環素類(tetracyclines):競爭 tRNA 與 A 位結合,選擇性透過主動運輸進入原核細胞。
- 胺基糖苷類(aminoglycosides)(如慶大黴素、阿米卡星):導致 mRNA 錯讀,合成異常蛋白。
- 氯黴素(chloramphenicol):抑制轉肽作用(transpeptidation)。
- 紅黴素(erythromycin):抑制 tRNA 從 A 位移至 P 位的位移(translocation)。
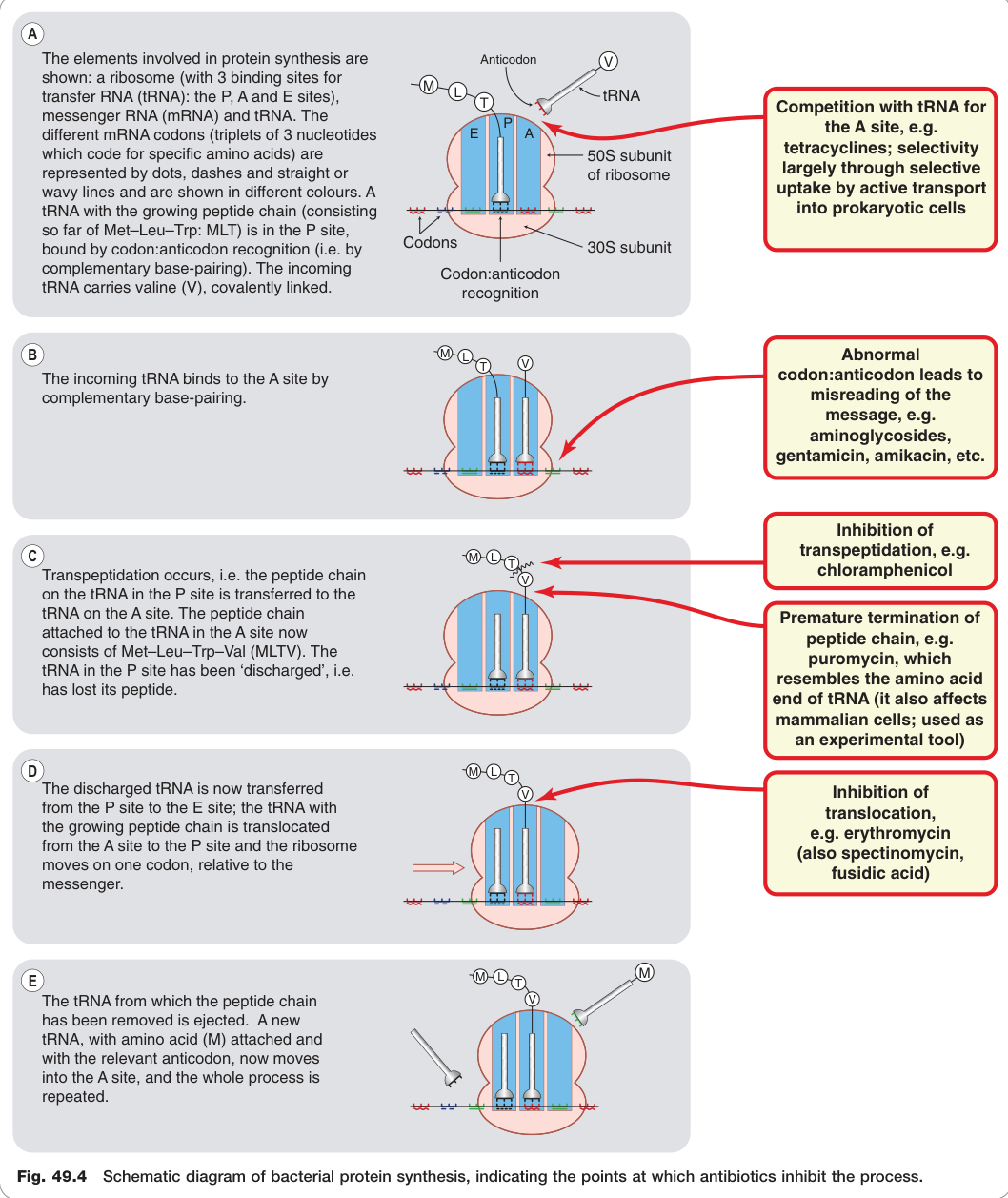
Figure 49.4:細菌蛋白質合成(核糖體轉譯)示意圖,標示各類抗生素(四環素類、胺基糖苷類、氯黴素、紅黴素等)的抑制作用位點
核酸合成#
可在五個層次干擾:
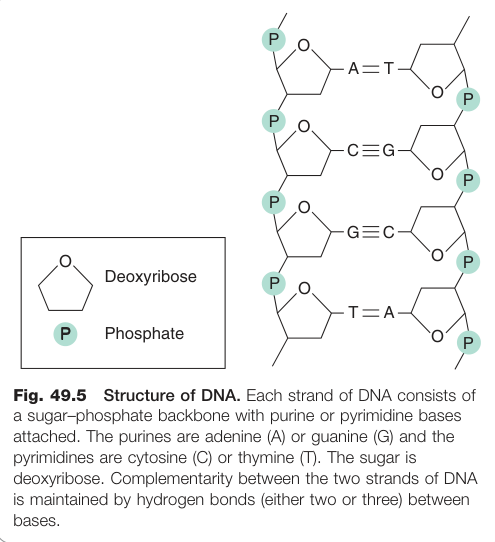
Figure 49.5:DNA 雙股結構示意圖,每條鏈由醣磷骨架與嘌呤(腺嘌呤 A、鳥嘌呤 G)及嘧啶(胞嘧啶 C、胸腺嘧啶 T)鹼基組成
- 抑制核苷酸合成(見 Class II)
- 改變 DNA 模板的鹼基配對特性:吖啶類嵌入 DNA,造成移碼突變(frameshift mutation)。

Figure 49.7:移碼突變(frameshift mutation)對 RNA 及蛋白質合成的影響範例——鹼基缺失或插入導致讀碼框位移,造成後續胺基酸序列全部錯誤
- 抑制 DNA 或 RNA 聚合酶:
- 利福平(rifampicin)/ 利福黴素(rifamycin):結合細菌 RNA 聚合酶,特異性高,用於結核病治療。
- 阿昔洛韋(aciclovir):在疱疹病毒感染細胞中被病毒激酶磷酸化,抑制病毒 DNA 聚合酶。
- 齊多夫定(zidovudine)/ 地達諾辛(didanosine):競爭逆轉錄酶(reverse transcriptase),用於 RNA 反轉錄病毒。
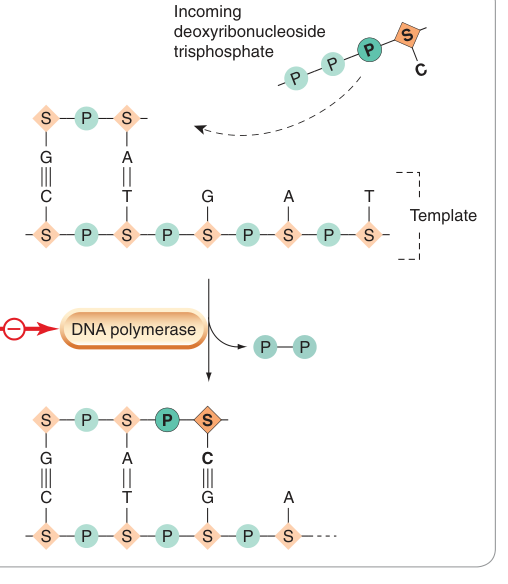
Figure 49.8:DNA 複製機制示意圖,標示以 DNA 聚合酶為靶點的各類抗生素抑制位點
- 抑制 DNA 旋轉酶(DNA gyrase):
- 氟喹諾酮類(fluoroquinolones)(如環丙沙星 ciprofloxacin):抑制細菌 DNA 旋轉酶(拓撲異構酶 II),對革蘭氏陰性菌特別有效;哺乳動物拓撲異構酶 II 對此類藥物不敏感。
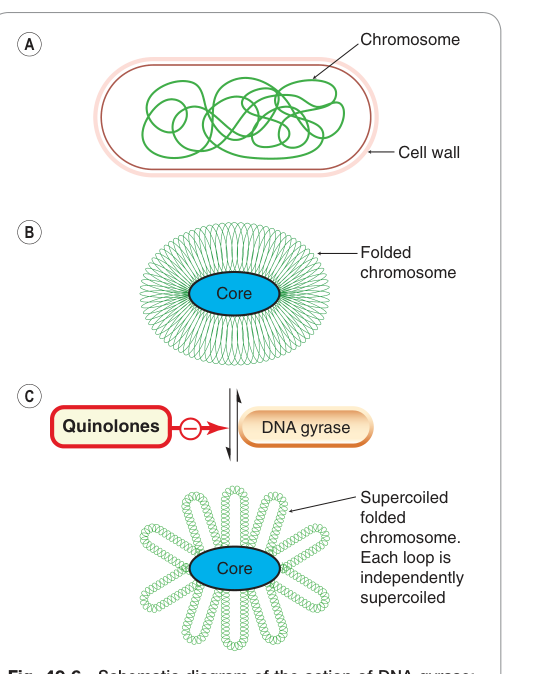
Figure 49.6:DNA gyrase(拓撲異構酶 II)的作用機制示意圖,及 quinolones 的作用位點
- 直接作用於 DNA:烷化劑(氮芥衍生物、亞硝脲類)僅用於癌症化療。
細胞形成結構作為靶點#
細胞膜#
- 多黏菌素類(polymyxins):陽離子肽抗生素,以去污劑方式破壞細菌磷脂膜。
- 多烯類(polyenes)(如制黴菌素 nystatin、兩性黴素 amphotericin):與真菌細胞膜特有的**麥角固醇(ergosterol)**結合,作為離子載體造成陽離子滲漏。
- 唑類(azoles)(如伊曲康唑 itraconazole):抑制麥角固醇合成,破壞真菌膜功能。
細胞內胞器#
- 苯並咪唑類(benzimidazoles)(如阿苯達唑 albendazole):選擇性結合寄生蟲微管蛋白,阻止微管形成(驅蟲藥)。
- 長春花生物鹼(vinca alkaloids)(長春鹼 vinblastine、長春新鹼 vincristine):干擾有絲分裂微管,用於抗癌。
- 氯喹(chloroquine):抑制瘧原蟲食物液泡中的血紅素聚合酶,阻止血紅素解毒。
肌肉纖維#
- 哌嗪(piperazine):激動線蟲 GABA 門控氯離子通道,使肌肉超極化而麻痺蟲體。
- 阿維菌素類(avermectins):增加線蟲肌肉 Cl⁻ 滲透性,造成麻痺。
抗菌藥物耐藥性#
自 1940 年代起,有效抗菌藥的開發伴隨著耐藥菌株的持續湧現。目前部分金黃色葡萄球菌及腸球菌株對幾乎所有現有抗生素均耐藥,形成嚴重的臨床威脅。
耐藥性的傳播途徑#
- 細菌在人際間傳播(帶著耐藥基因的細菌本身擴散)
- 耐藥基因在細菌間傳播(通常藉由質粒)
- 耐藥基因在菌體內不同遺傳元件間轉移(藉由轉座子)
遺傳決定因素#
染色體突變#
- 自發突變率約為 1/10⁷,但大量細菌使得耐藥突變株出現機率相當高。
- 重要實例:耐甲氧西林金黃色葡萄球菌(MRSA)、耐藥結核分枝桿菌。
基因擴增#
- 抗生素暴露可誘導既有耐藥基因(如藥物水解酶、外排泵基因)拷貝數增加。
質粒(Plasmids)#
- 質粒是細胞質中可獨立複製的環狀染色體外 DNA。
- 攜帶耐藥基因(r genes)的稱為 R 質粒,是臨床耐藥最常見的決定因素。
- 接合性質粒(conjugative plasmids)可透過接合(conjugation)——細胞間形成性菌毛(sex pili)管道——在同種或不同種細菌間轉移。
轉座子(Transposons)與整合子(Integrons)#
- 轉座子:可在質粒間、質粒與染色體間轉移的 DNA 片段,可攜帶一個或多個耐藥基因並在複製過程中使供體與受體雙方均獲得耐藥基因。
- 整合子:可整合多個**基因卡匣(gene cassettes)**的移動元件,形成多重耐藥陣列,使多重耐藥在細菌間極為高效地傳播。
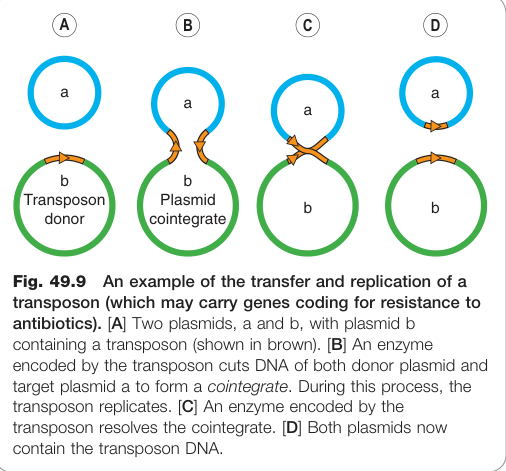
Figure 49.9:轉座子(transposon)在質粒間轉移與複製的機制示意圖——轉座後供體與受體質粒均攜帶耐藥基因,加速多重耐藥傳播
耐藥的生化機制#
| 機制 | 代表藥物/例子 |
|---|---|
| 酶滅活藥物 | β-內醯胺酶水解青黴素;乙醯轉移酶滅活氯黴素;激酶類滅活胺基糖苷類 |
| 藥物結合位點改變 | 核糖體 30S 或 50S 亞基突變(胺基糖苷類、紅黴素);DNA 旋轉酶突變(氟喹諾酮類);RNA 聚合酶突變(利福平) |
| 藥物累積減少 | 質粒介導外排泵主動泵出四環素;外膜孔蛋白(porins)減少阻礙親水性抗生素進入 |
| 旁路替代途徑 | 對甲氧苄啶耐藥的 DHFR 低親和力異構體;對磺胺耐藥的低親和力二氫蝶呤合成酶 |
多重耐藥現況#
MRSA 已成為院內感染的主要威脅,曾是最後防線的萬古黴素(vancomycin),自 1997 年起已出現敏感性降低的菌株。耐萬古黴素腸球菌(VRE)亦迅速增加,且存在耐藥基因轉移至金黃色葡萄球菌的風險。
結核分枝桿菌(Mycobacterium tuberculosis)亦出現能規避幾乎所有抗結核藥物的菌株,目前結核病死亡人數已超過瘧疾與愛滋病的總和。
耐藥性加劇的原因不僅是細菌的適應:
- 人類及獸醫用藥的濫用,促進了耐藥菌株的選擇。
- 製藥業對新型抗生素研發的投入曾大幅縮減(1967 年美國外科醫師總監宣告傳染病已被征服後),直至近年才恢復。
- 大多數現有抗生素僅是少數基本骨架(如 β-內醯胺核)的漸進式改良,突破性新結構匱乏。