概覽#
人體每天製造約 100 億個新細胞,同時也必須清除相同數量的舊細胞,以維持組織穩定。本章涵蓋四大主題:
- 細胞增殖(cell proliferation):細胞週期的調控機制
- 細胞凋亡(apoptosis):程序性細胞死亡的路徑與意義
- 修復與癒合(repair and healing):損傷後組織的重建過程
- 再生(regeneration):幹細胞與組織再生的治療潛力
這些過程皆與生長因子、胞外基質及藥物作用密切相關,也是腫瘤藥理學、抗發炎藥物與再生醫學的重要理論基礎。
細胞增殖#
細胞週期#
細胞增殖涉及多種生理與病理過程,包括生長、傷口癒合、肥大、增生及腫瘤形成。增殖細胞經歷一套稱為**細胞週期(cell cycle)**的有序事件:
| 時期 | 說明 |
|---|---|
| G1 | 為 DNA 合成做準備 |
| S | DNA 合成與染色體複製 |
| G2 | 為分裂做準備 |
| M(有絲分裂) | 分裂為兩個子細胞 |
靜止細胞(G0 期)受生長因子刺激後,可被驅入 G1 期。神經元與骨骼肌細胞終生停留在 G0;骨髓細胞與腸道上皮細胞則每日分裂。
週期中有兩個檢查點(check points),分別位於 S 期與 M 期入口。DNA 受損時,週期會在這些檢查點停止以利修復;若檢查點功能失常,是癌症形成的重要特徵。
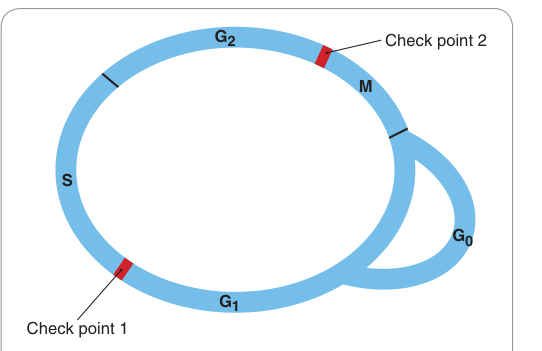
Figure 5.1:增殖細胞的細胞週期各主要時期
正性調節因子:Cyclin/CDK 複合體#
**細胞週期依賴激酶(cyclin-dependent kinases, cdks)是推動週期前進的核心酵素。每個 cdk 必須與對應的週期蛋白(cyclin)結合才會活化,活化後對特定蛋白質進行磷酸化;磷酸化完成後,cyclin 透過泛素/蛋白酶體(ubiquitin/proteasome)**系統被降解。
四種主要複合體及其作用:
- Cyclin D / cdk 4 & 6:G1 期啟動;磷酸化並釋放 Rb 蛋白的抑制
- Cyclin E / cdk 2:G1 → S 期轉換(通過檢查點 1)
- Cyclin A / cdk 1 & 2:S 期至 G2 期推進
- Cyclin B / cdk 1:G2 → M 期轉換(通過檢查點 2)
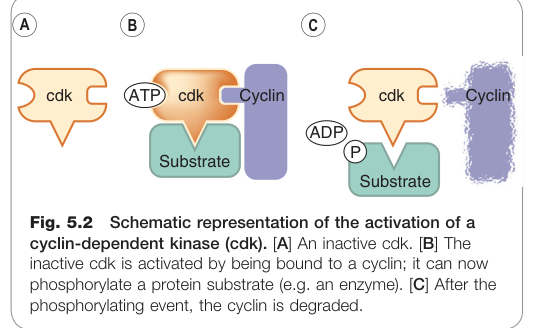
Figure 5.2:cyclin-dependent kinase(cdk)活化示意圖——非活性 cdk 與 cyclin 結合後才具有磷酸化受質的能力
有絲分裂(M 期)分為四個階段:
- 前期(prophase):染色體凝縮,核膜崩解
- 中期(metaphase):染色體排列於赤道板
- 後期(anaphase):染色分體被拉向兩極
- 末期(telophase):核膜重建,細胞質分裂形成兩個子細胞
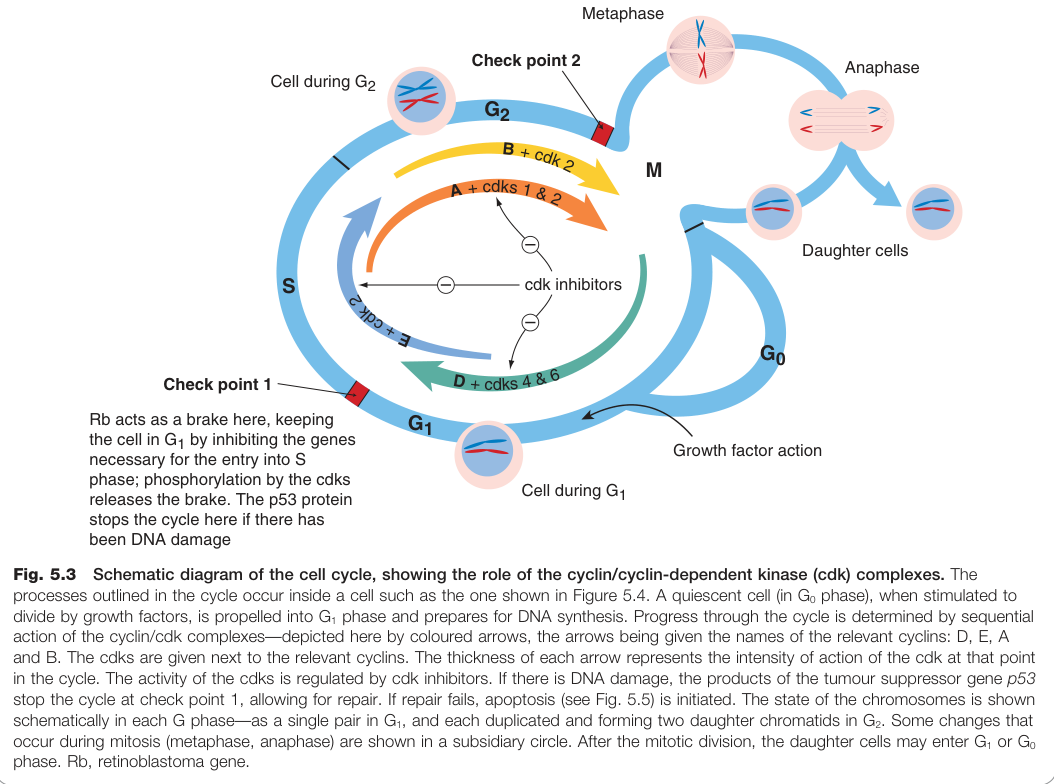
Figure 5.3:細胞週期示意圖——各時期中 cyclin/cdk 複合體的角色與兩個檢查點的位置
負性調節因子#
兩大家族的 cdk 抑制蛋白在檢查點 1 發揮主要作用:
- CIP/KIP 家族:p21、p27、p57
- Ink 家族:p16、p19、p15
p53 基因被稱為「基因組守護者」。正常細胞中 p53 蛋白濃度很低;DNA 受損時,p53 蛋白累積並活化 p21 的轉錄,p21 再抑制 cyclin/cdk 複合體,使週期在檢查點 1 停止以供修復。若修復失敗,p53 則觸發凋亡。
若檢查點功能失常(如 p53 突變),DNA 受損的細胞得以繼續增殖,是癌症發生的核心機制之一。
細胞、生長因子與胞外基質的互動#
生長因子#
生長因子經由**受體酪氨酸激酶(receptor tyrosine kinases)**或受體偶聯激酶啟動下游訊號,最終活化控制細胞週期的基因。重要的生長因子包括:
- 纖維母細胞生長因子(fibroblast growth factor, FGF)
- 表皮生長因子(epidermal growth factor, EGF)
- 血小板衍生生長因子(platelet-derived growth factor, PDGF)
- 血管內皮生長因子(vascular endothelial growth factor, VEGF)
- 轉化生長因子-β(transforming growth factor-β, TGF-β)
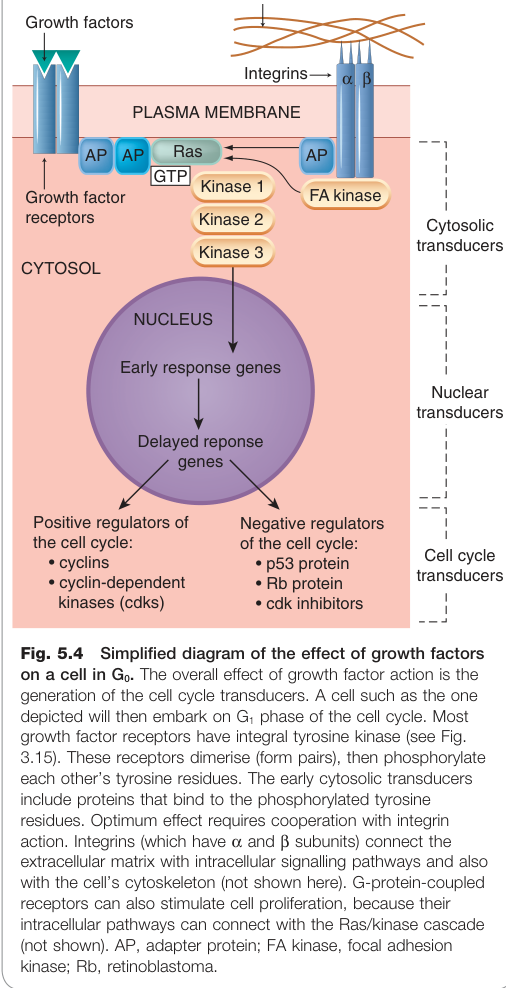
Figure 5.4:生長因子對 G0 期細胞的作用——最終活化控制細胞週期的基因,驅動細胞進入增殖
胞外基質(ECM)#
**胞外基質(extracellular matrix, ECM)**由細胞本身分泌,主要成分包括:
- 纖維形成元素:膠原蛋白(collagen,ECM 的主要蛋白質)、彈性蛋白(elastin)
- 非纖維元素:蛋白聚糖(proteoglycans)、黏附蛋白(如纖連蛋白 fibronectin)
ECM 不僅提供支撐框架,也透過細胞表面的**整合素(integrins)**深刻影響細胞行為,並充當生長因子的儲存庫。
整合素#
整合素是跨膜受體(具 α 和 β 亞基),與 ECM 元素結合後:
- 透過焦點黏附激酶(focal adhesion kinase)活化激酶級聯
- 與生長因子訊號路徑產生交互作用,是最佳細胞增殖所必需的
- 抑制 cdk 抑制蛋白、促進 cyclin A 和 D 的表現
- 刺激抗凋亡訊號
基質金屬蛋白酶(MMPs)#
**基質金屬蛋白酶(matrix metalloproteinases, MMPs)**由局部細胞分泌(初始為無活性前驅體),在組織生長、修復與重塑時降解 ECM。其功能包括:
- 為增殖所需的細胞數增加創造局部空間
- 從 ECM 釋放被隔離的生長因子
- 將某些生長因子從前驅形式活化為活性形式
MMPs 的活性受**TIMPS(組織金屬蛋白酶抑制劑)**調控。過度的 MMP 活性參與了類風濕性關節炎、腫瘤侵襲與轉移等病理過程。
血管新生(Angiogenesis)#
血管新生是由現有微血管形成新毛細血管的過程,細胞增殖時必然伴隨發生。VEGF 是最重要的促血管新生細胞因子。
步驟:
- 蛋白酶局部降解基底膜
- 內皮細胞遷移,形成新芽
- 後繼內皮細胞在 VEGF 的影響下增殖
- 新毛細血管周圍沉積基質
針對 VEGF 的單株抗體**貝伐珠單抗(bevacizumab)**已用於多種癌症的輔助治療,也可注射入眼內治療年齡相關黃斑部病變。
細胞凋亡#
細胞凋亡是一種由基因程式化的自我毀滅機制,透過一系列生化事件有序執行,與壞死(necrosis)截然不同——後者是細胞受損後的無序解體,並觸發發炎反應。
生理與病理意義#
生理功能:
- 胚胎發育期塑造器官形態
- 每日清除 100 億個時效過期的細胞(如腸道黏膜、中性球)
- 建立免疫系統的自我耐受性
- 清除 DNA 異常細胞,防止惡性轉化
過度凋亡相關疾病:
- 阿茲海默症、多發性硬化症、帕金森氏症等神經退化疾病
- 心肌梗塞、中風、脊髓損傷等急性組織損傷
- HIV 感染造成的 T 細胞耗竭
凋亡不足相關疾病:
- 癌細胞逃避免疫清除、化療抗藥性
- 自體免疫疾病(如重症肌無力、類風濕性關節炎、氣喘)
- 病毒感染細胞未被有效清除
凋亡是預設反應。細胞存活需要特定存活因子(生長因子、細胞激素、荷爾蒙、細胞間接觸訊號)持續主動地抑制自毀機制;一旦存活因子撤離(「被遺忘的死亡,death by neglect」),凋亡便自動啟動。
形態學變化#
凋亡細胞的特徵:
- 細胞圓縮,染色質凝縮成密塊
- 細胞質縮小,質膜形成出芽(blebbing)
- 最終由胱天蛋白酶(caspases)將細胞分解為膜包被的小體(membrane-bound entities)
- 這些小體顯示「吃我」訊號(如磷脂醯絲胺酸外露),被巨噬細胞吞噬清除
主要分子機制#
Caspases#
**胱天蛋白酶(caspases)**是一家族的半胱胺酸蛋白酶,在細胞中以無活性形式存在。
- 啟動型 caspases(initiator caspases):接收凋亡訊號(如 caspase 8、caspase 9)
- 效應型 caspases(effector caspases):執行細胞死亡(如 caspase 3),切割並破壞 DNA 修復酶、蛋白激酶 C、細胞骨架等,並活化 DNAase 將基因組 DNA 切成約 180 bp 的片段
Bcl-2 家族#
Bcl-2 家族蛋白在粒線體表面競爭調控凋亡命運:
- 促凋亡成員:Bax、Bak、Bid——促進粒線體釋放細胞色素 c(cytochrome c)
- 抗凋亡成員:Bcl-2 本身——抑制細胞色素 c 的釋放
兩條主要凋亡路徑#
死亡受體路徑#
- 細胞表面有腫瘤壞死因子受體(TNFR)超家族成員,即死亡受體(death receptors)(如 Fas/CD95、TNFR-1)
- 外部配體(如 TNF、TRAIL)結合後,受體三聚化,招募接合蛋白
- 活化啟動型 caspase 8,再啟動效應型 caspases
粒線體路徑#
- 由 DNA 損傷或存活因子撤離觸發
- p53 蛋白活化促凋亡 Bcl-2 成員(Bid、Bax、Bak)
- 粒線體釋放細胞色素 c,與 Apaf-1 及前 caspase 9 組成凋亡小體(apoptosome)
- 凋亡小體活化 caspase 9,進一步啟動效應型 caspases
兩條路徑最終匯聚於效應型 caspase(如 caspase 3)。死亡受體路徑中的 caspase 8 也可活化促凋亡 Bcl-2,從而交叉啟動粒線體路徑。
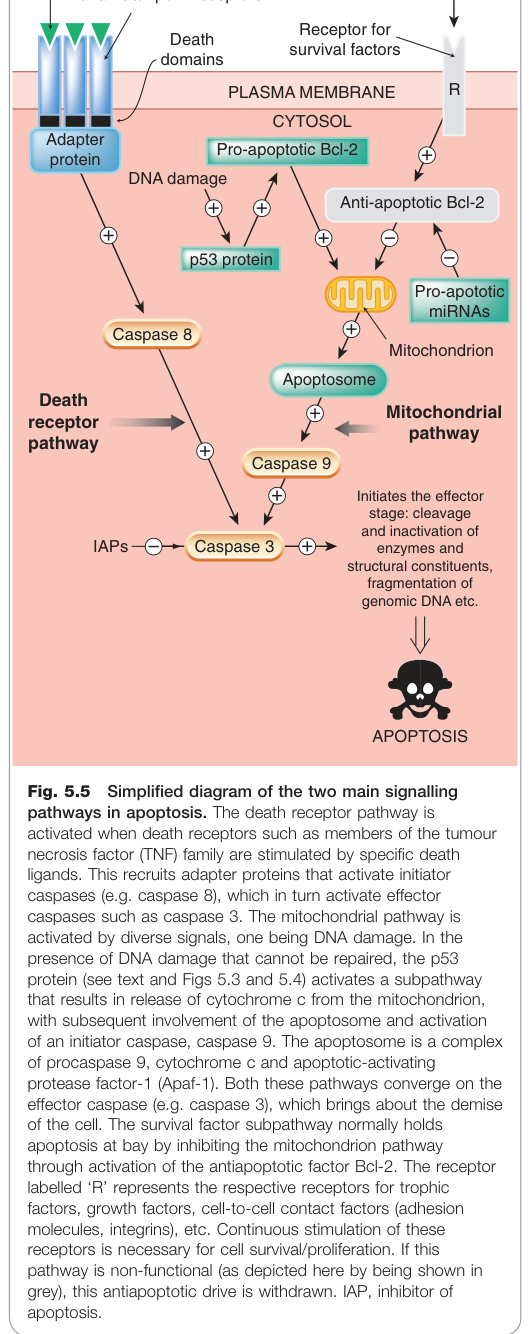
Figure 5.5:凋亡兩大主要訊號路徑——死亡受體路徑(TNF 家族)與粒線體路徑,最終匯聚於 caspase 級聯
MicroRNA 與細胞週期/凋亡#
**微型 RNA(microRNAs, miRNAs)**是一類小型非編碼 RNA,可抑制控制細胞週期、凋亡、分化與發育的基因表現。約 3% 的人類基因編碼 miRNA,估計多達 30% 的蛋白質編碼基因受 miRNA 調控。miRNA 的異常表現與癌症、糖尿病、肥胖、神經退化疾病等多種疾病相關,也是新藥開發的潛在靶點。
修復、癒合與再生#
修復與癒合#
損傷或組織缺失後,修復過程依序涉及:
- 細胞遷移與血管新生
- 結締組織細胞增殖
- 胞外基質合成
- 組織重塑
TGF-β 是多個修復步驟的關鍵細胞激素。發炎反應與修復過程在細胞與機制上有大量重疊。
增生(Hyperplasia)#
細胞增殖與基質擴張是慢性發炎、超敏反應及自體免疫疾病(如類風濕性關節炎、乾癬、慢性氣喘、腎小球腎炎)的重要特徵。
幹細胞與再生#
**再生(regeneration)**意味著受損組織的完整復原,有別於修復後留下瘢痕的癒合。
- 兩棲類等低等動物擁有強大的再生能力,因為其器官含有豐富的幹細胞(stem cells),且分化細胞可去分化成幹細胞
- 哺乳動物在演化中大多喪失了這種能力,僅血液細胞、腸道上皮和皮膚外層能持續更新
- 肝臟是哺乳動物中罕見的例外,切除至少 75% 後仍可在數週內再生
目前正在研究的幹細胞來源包括:
- 胚胎幹細胞(embryonic stem cells)——倫理議題限制應用
- 骨髓來源間葉幹細胞(bone marrow-derived mesenchymal stem cells)
- 肌肉來源幹細胞(muscle-derived stem cells)
- 人類誘導多能幹細胞(human-induced pluripotent stem cells, iPSC)
- 組織常駐前驅細胞(tissue-residing progenitor cells)
修復可能是哺乳動物在演化上以快速封閉缺損交換再生能力的折衷結果。快速修復對存活更具短期優勢。
治療前景#
調控凋亡的藥物#
促凋亡策略(主要針對癌症):
- 奧利默森(oblimersen):Bcl-2 的反義化合物,針對慢性淋巴性白血病進行第三期試驗
- 奧巴托拉(obatoclax):Bcl-2 抑制小分子,針對血液腫瘤進行 I/II 期試驗
- TRAIL 激動抗體(mapatumumab、lexatumumab):針對實體瘤和淋巴瘤的 I/II 期試驗
- 硼替佐米(bortezomib):蛋白酶體抑制劑,抑制 Bcl-2 抗凋亡蛋白的降解,現已用於多發性骨髓瘤
- survivin 抑制劑:survivin 是高度腫瘤特異性的內源性 caspase 抑制劑,其抑制劑正在臨床試驗中
抗凋亡策略(針對退化性疾病):目前臨床試驗結果尚不理想,多種化合物被發現缺乏療效。
調控血管新生的藥物#
- 尋找有效 MMP 抑制劑的工作持續進行,但臨床試驗效果有限
- 貝伐珠單抗(bevacizumab)(抗 VEGF 單株抗體)已核准用於多種癌症治療
調控細胞週期的藥物#
- CDK 抑制劑(如 flavopiridol):靶向 ATP 結合位點,抑制所有 cdks,並兼具促凋亡與抗血管新生效果
- 受體酪氨酸激酶抑制劑(如 imatinib、gefitinib、erlotinib):已用於癌症治療
- 硼替佐米:同時抑制蛋白酶體以防止促凋亡蛋白降解
理論上,細胞增殖、凋亡、ECM 重塑和血管新生等所有過程都可作為新藥開發的靶點,預期在未來十年間會有更多藥物問世,尤其在腫瘤藥理學領域。