本章聚焦於藥物在細胞層次產生效應的短時程機制(毫秒至數小時),主要涵蓋興奮(excitation)、收縮(contraction)與分泌(secretion)。這些機制幾乎都由細胞內游離鈣離子濃度([Ca²⁺]i)所調控。
鈣離子的細胞內調控#
為何 Ca²⁺ 如此重要#
- 靜息細胞的 [Ca²⁺]i 維持在約 10⁻⁷ M(0.1 µmol/l)
- 細胞外液 [Ca²⁺]o 約 2.4 mM,兩側存在巨大的濃度梯度
- Ca²⁺ 是酵素、收縮蛋白、囊泡蛋白等功能蛋白的關鍵調節者
維持低 [Ca²⁺]i 需要:(a) 透過漿膜主動輸出 Ca²⁺;(b) 將 Ca²⁺ 泵入內質網(ER);(c) 漿膜與 ER 膜對 Ca²⁺ 的低通透性。
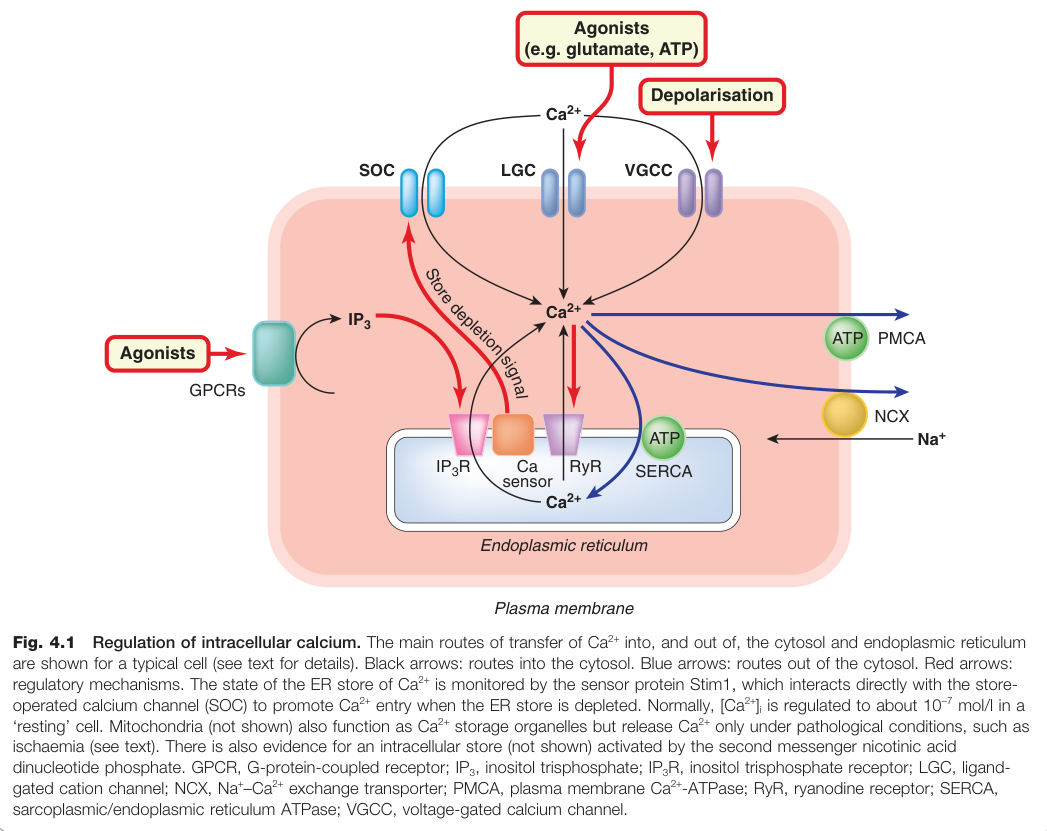
Figure 4.1:細胞內 Ca²⁺ 調控的主要路徑,包括 Ca²⁺ 進出細胞質與內質網的轉運機制
Ca²⁺ 進入細胞的途徑#
Ca²⁺ 通過以下四種主要途徑進入細胞:
- 電壓閘控鈣離子通道(voltage-gated calcium channels, VGCC)
- 配體閘控鈣離子通道(ligand-gated calcium channels, LGC)
- 儲存操控鈣離子通道(store-operated calcium channels, SOC)
- Na⁺–Ca²⁺ 交換子(Na⁺–Ca²⁺ exchanger, NCX)(雙向)
電壓閘控鈣離子通道#
依據活化/去活化動力學、電壓閾值、電導及藥物敏感性,分為五種亞型:
| 亞型 | 主要分布 | 代表藥物 |
|---|---|---|
| L 型 | 心肌與平滑肌(收縮主要 Ca²⁺ 來源) | 二氫吡啶類(nifedipine)、維拉帕米、地爾硫卓(阻斷);BayK 8644(活化) |
| N 型 | 神經末梢(主要傳遞物釋放) | ω-conotoxin(芋螺毒素)阻斷;ziconotide 用於鎮痛 |
| T 型 | 心臟節律細胞、神經元 | mibefradil 阻斷 |
| P/Q 型 | 神經末梢(傳遞物釋放) | ω-agatoxin(蜘蛛毒)阻斷 |
| R 型 | 神經元樹突 | — |
許多藥物透過 G 蛋白偶合受體(GPCR)間接調控鈣離子通道;直接作用於 VGCC 的臨床藥物以「鈣離子拮抗劑(Ca²⁺ antagonists)」為代表。
配體閘控通道#
- 大多數興奮性神經傳遞物所開啟的陽離子通道對 Ca²⁺ 有通透性
- **NMDA 型麩胺酸受體(NMDA receptor)**對 Ca²⁺ 通透性特別高,為中樞神經元 Ca²⁺ 進入的主要途徑之一
- 過度激活 NMDA 受體造成大量 Ca²⁺ 進入,可引發興奮毒性(excitotoxicity),與多種神經退化疾病有關
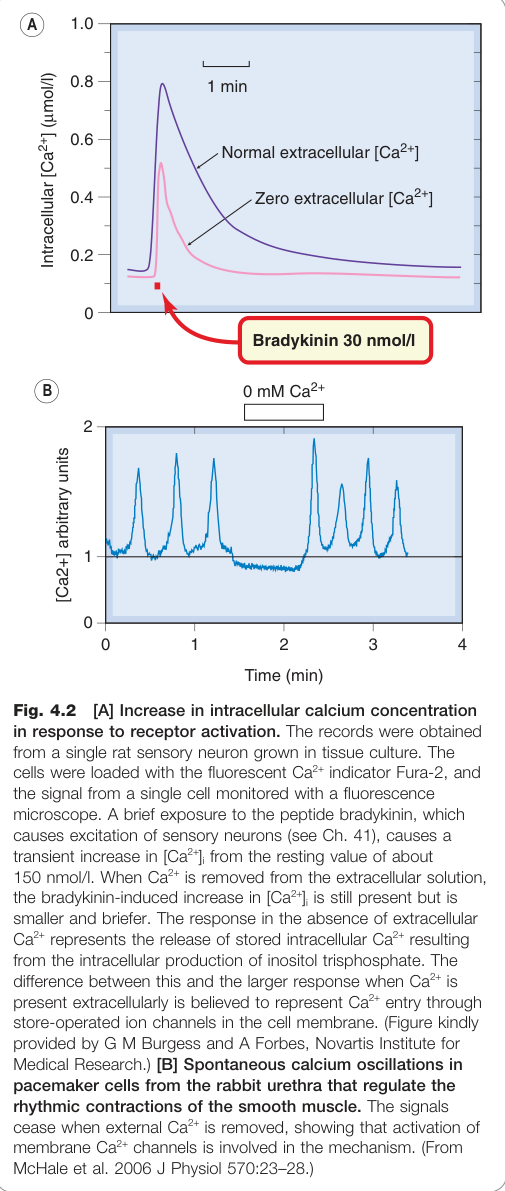
Figure 4.2:受體活化後細胞內 Ca²⁺ 濃度的上升,以及螢光指示劑記錄的振盪波形
儲存操控鈣離子通道#
- SOC 位於漿膜,當 ER 的 Ca²⁺ 儲存耗盡時才開放
- 感測蛋白 Stim1 監測 ER 的 Ca²⁺ 含量,直接與漿膜的通道蛋白 Orai1 交互作用,促進 Ca²⁺ 進入
Ca²⁺ 排出與儲存交換#
- 漿膜 Ca²⁺-ATPase(PMCA):主動將 Ca²⁺ 泵出細胞
- SERCA(肌漿/內質網 Ca²⁺-ATPase):將 Ca²⁺ 泵入 ER/SR;**毛剪菜素(thapsigargin)**可特異性阻斷此泵(實驗工具,無治療意義)
- Na⁺–Ca²⁺ 交換子:每次轉運 3 個 Na⁺ 換 1 個 Ca²⁺,能量來自 Na⁺ 電化學梯度;**地高辛(digoxin)**透過抑制 Na⁺ 輸出間接升高 [Ca²⁺]i,增強心肌收縮力
Ca²⁺ 從 ER/SR 釋出#
ER/SR 膜上有兩種主要 Ca²⁺ 釋放通道:
- IP₃ 受體(IP₃R):由第二訊息 IP₃(inositol trisphosphate)激活,是 GPCR 活化後升高 [Ca²⁺]i 的主要機制
- 雷諾丁受體(ryanodine receptor, RyR):
- 骨骼肌中由 T 管的二氫吡啶受體直接偶聯觸發
- 其他細胞中由輕微的 [Ca²⁺]i 上升即可觸發,產生鈣誘導鈣釋放(Ca²⁺-induced Ca²⁺ release, CICR),形成放大效應
- 咖啡因可增加 RyR 對 Ca²⁺ 的敏感性(高濃度時才明顯)
- 丹特羅林(dantrolene)阻斷 RyR,用於治療惡性高熱(malignant hyperthermia)
Ca²⁺ 訊號具有時間與空間的複雜模式(振盪、波)。正回饋(CICR)和多種負回饋機制共同決定 Ca²⁺ 訊號的波形,這是平滑肌和神經細胞自發節律性活動的基礎。
鈣調蛋白(Calmodulin)#
- Ca²⁺ 透過與**鈣調蛋白(calmodulin)**結合發揮功能
- 鈣調蛋白有四個 Ca²⁺ 結合位點,全佔據後發生構象變化,暴露疏水性結構域,進而調節至少 40 種功能蛋白(包括激酶、磷酸酶、通道蛋白等)
興奮性——動作電位#
興奮性(excitability)指細胞對膜去極化產生全有全無(all-or-nothing)電位反應的能力,即動作電位(action potential),見於神經元、骨骼肌、心肌、平滑肌及部分內分泌腺細胞。
靜息細胞的離子平衡#
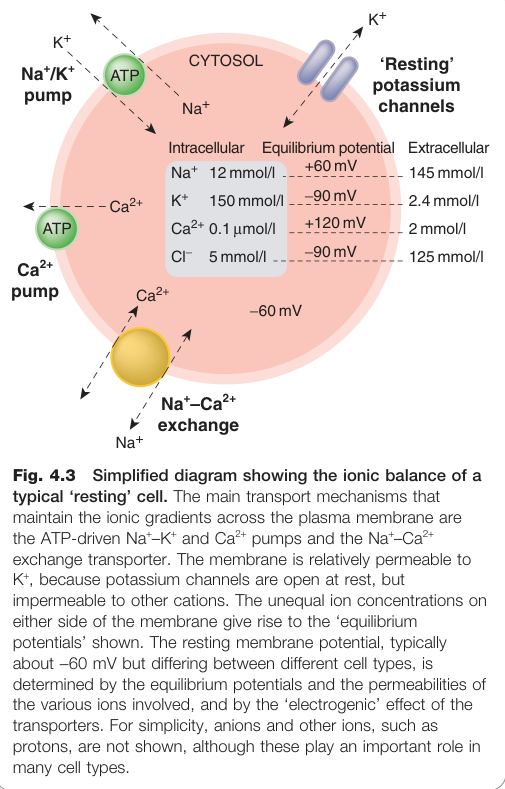
Figure 4.3:典型靜息細胞的離子平衡,顯示維持漿膜離子梯度的 Na⁺–K⁺-ATPase 等主要轉運機制
- 細胞膜電位在 −30 至 −80 mV(依細胞種類而異)
- Na⁺–K⁺-ATPase 主動輸出 Na⁺ / 輸入 K⁺,使細胞內 [K⁺]i 高、[Na⁺]i 低
- 靜息時膜對 K⁺ 通透性較高,膜電位約等於 K⁺ 平衡電位(−90 mV)
動作電位的離子基礎#
霍奇金(Hodgkin)、赫胥黎(Huxley)與卡茨(Katz)的經典研究揭示動作電位由兩個過程產生:
- Na⁺ 通透性快速上升(超過 −50 mV 時激活)→ 內向去極化電流 → 正回饋產生上升相
- K⁺ 通透性延遲上升(動力學較慢、不去活化)→ 外向電流 → 協助復極化
電壓閘控 Ca²⁺ 通道功能與 Na⁺ 通道類似,在心肌、平滑肌及神經分泌細胞中扮演動作電位產生及胞內訊號的重要角色。
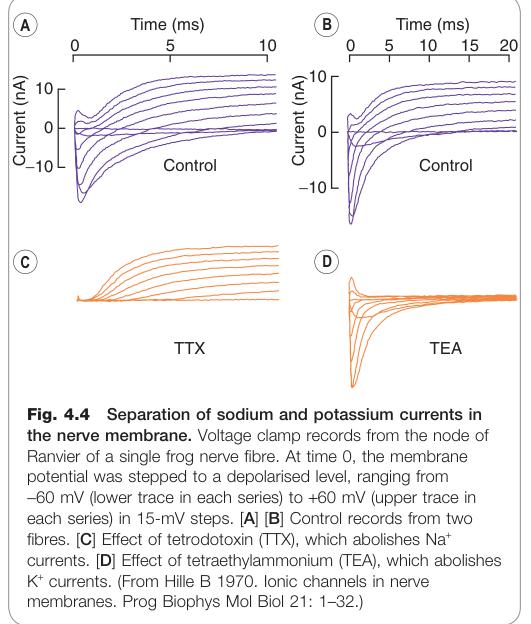
Figure 4.4:電壓箝制下青蛙神經纖維 Na⁺ 與 K⁺ 電流的分離記錄
通道的功能狀態與藥物的依賴性#
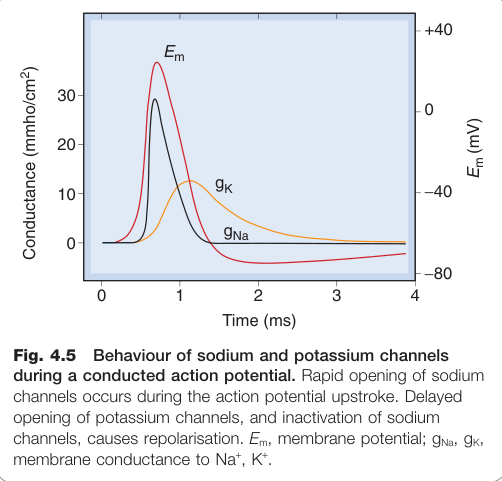
Figure 4.5:動作電位傳導期間 Na⁺ 通道快速開放(上升相)與 K⁺ 通道延遲開放(復極化)的行為
電壓閘控通道存在三種功能狀態:靜息(resting)、活化(activated,開放)、去活化(inactivated)。
- 使用依賴性(use dependence):對去活化態(inactivated state)親和力高的藥物,在高頻放電時阻斷效果更強,而不影響正常頻率的興奮性。抗心律不整藥及抗癲癇藥多具此特性。
- 電壓依賴性(voltage dependence):陽離子型 Na⁺ 通道阻斷藥在去極化時阻斷效果增強,在心律不整或癲癇灶中(細胞通常較去極化)可達到選擇性阻斷。
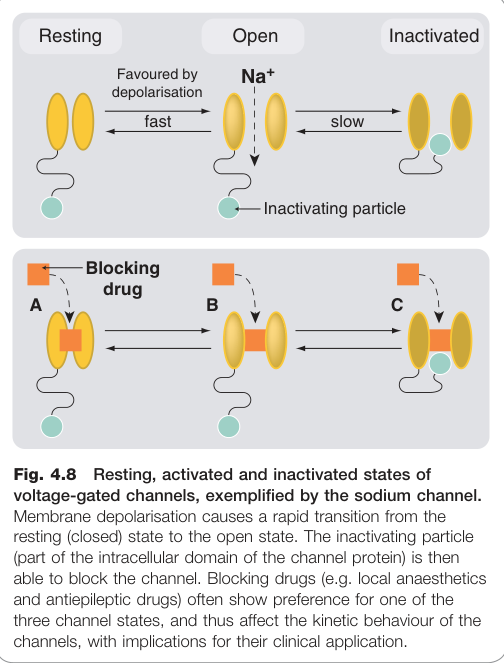
Figure 4.8:電壓閘控鈉離子通道的靜息、活化(開放)與去活化(失活)三態示意圖,以及藥物的作用位點
鈉離子通道#
- 結構含四個同源域,各有六個跨膜螺旋;S4 螺旋為電壓感測器
- 已鑑定九種分子亞型,心肌與感覺神經元的鈉離子通道對 河豚毒素(tetrodotoxin, TTX) 相對不敏感
- 河豚毒素(TTX)、蟾毒素(saxitoxin, STX):阻斷 Na⁺ 通道(實驗工具;局部麻醉藥機制相似)
- 藜蘆鹼(veratridine)、蟾皮毒素(batrachotoxin):造成 Na⁺ 通道持續活化 → 神經過度興奮
- 局部麻醉藥(Ch. 42)、抗心律不整藥(Ch. 21)、抗癲癇藥(Ch. 44):均以阻斷 Na⁺ 通道為主要機制
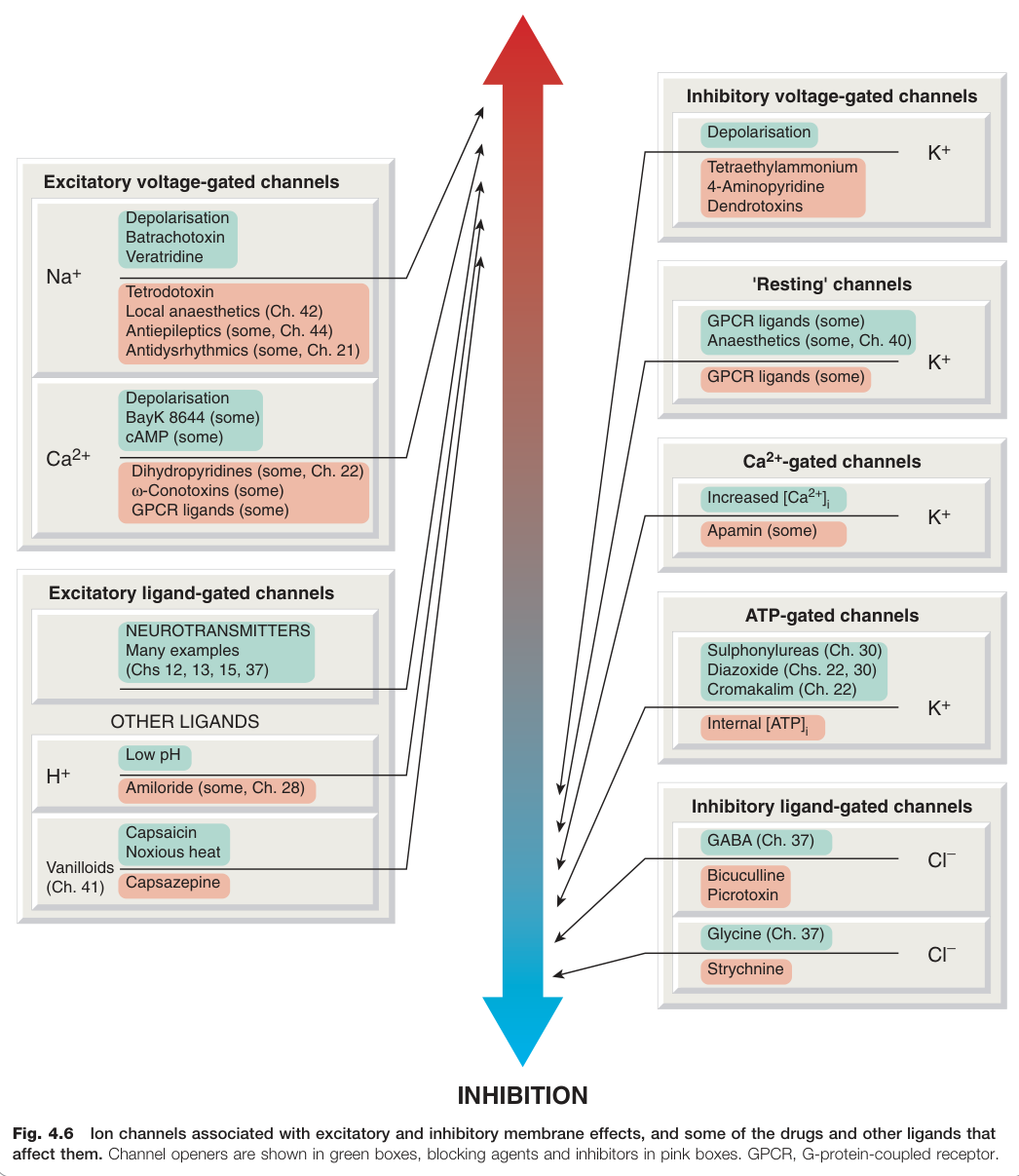
Figure 4.6:與興奮性和抑制性膜效應相關的離子通道,以及影響這些通道的藥物與配體
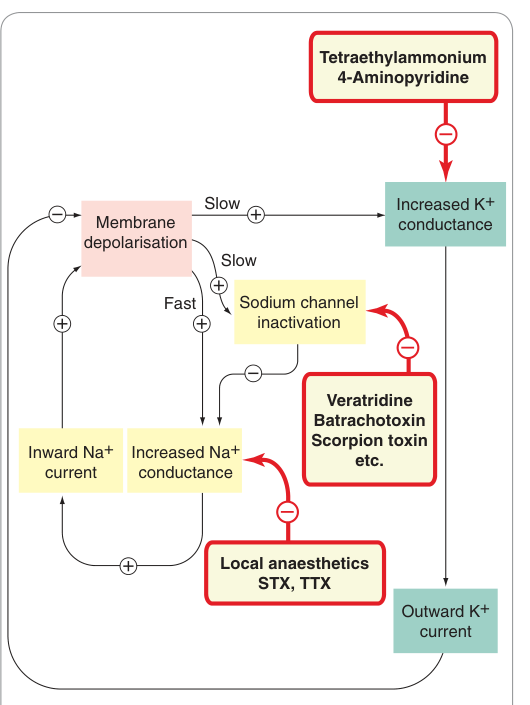
Figure 4.7:影響動作電位產生的通道的藥物與毒素作用位點,包括 Na⁺、K⁺、Ca²⁺ 通道;STX(saxitoxin)等毒素及局部麻醉藥均標示於此
鉀離子通道#
K⁺ 通道是調控細胞興奮性最多樣化的一族,分三大結構類別:
| 類別 | 代表亞型 | 功能 | 藥物相關性 |
|---|---|---|---|
| 電壓閘控(6T, 1P) | HERG、LQT | 動作電位復極化、限制最高放電頻率 | TEA、4-氨基吡啶阻斷;HERG 突變與 long QT 症候群相關 |
| 內向整流(2T, 1P) | G 蛋白活化型、ATP 敏感型 | GPCR 抑制性效應;控制胰島素分泌 | 磺醯脲類(glibenclamide)關閉 ATP 敏感型 K⁺ 通道;cromakalim 開啟 → 平滑肌鬆弛 |
| 雙孔域(4T, 2P) | TWIK、TREK、TASK | 維持靜息 K⁺ 電導,對抗去極化 | 某些亞型被揮發性麻醉劑(halothane)活化 |
肌肉收縮#
三種肌肉的收縮基礎相同——肌動蛋白(actin)與肌凝蛋白(myosin)在 Ca²⁺ 升高下的相互作用——但興奮–收縮偶聯(excitation–contraction coupling)的機制各異。
骨骼肌#
- 動作電位依賴 電壓閘控 Na⁺ 通道,由運動終板傳導至 T 管
- T 管膜上的 **L 型 Ca²⁺ 通道(二氫吡啶受體)**與 SR 的 RyR 直接機械偶聯
- 去極化 → RyR 開放 → Ca²⁺ 從 SR 快速釋出 → Ca²⁺ 與**肌鈣蛋白(troponin)**結合 → 解除肌動蛋白阻塞 → 收縮
- 反應快速且直接,較少受藥物影響
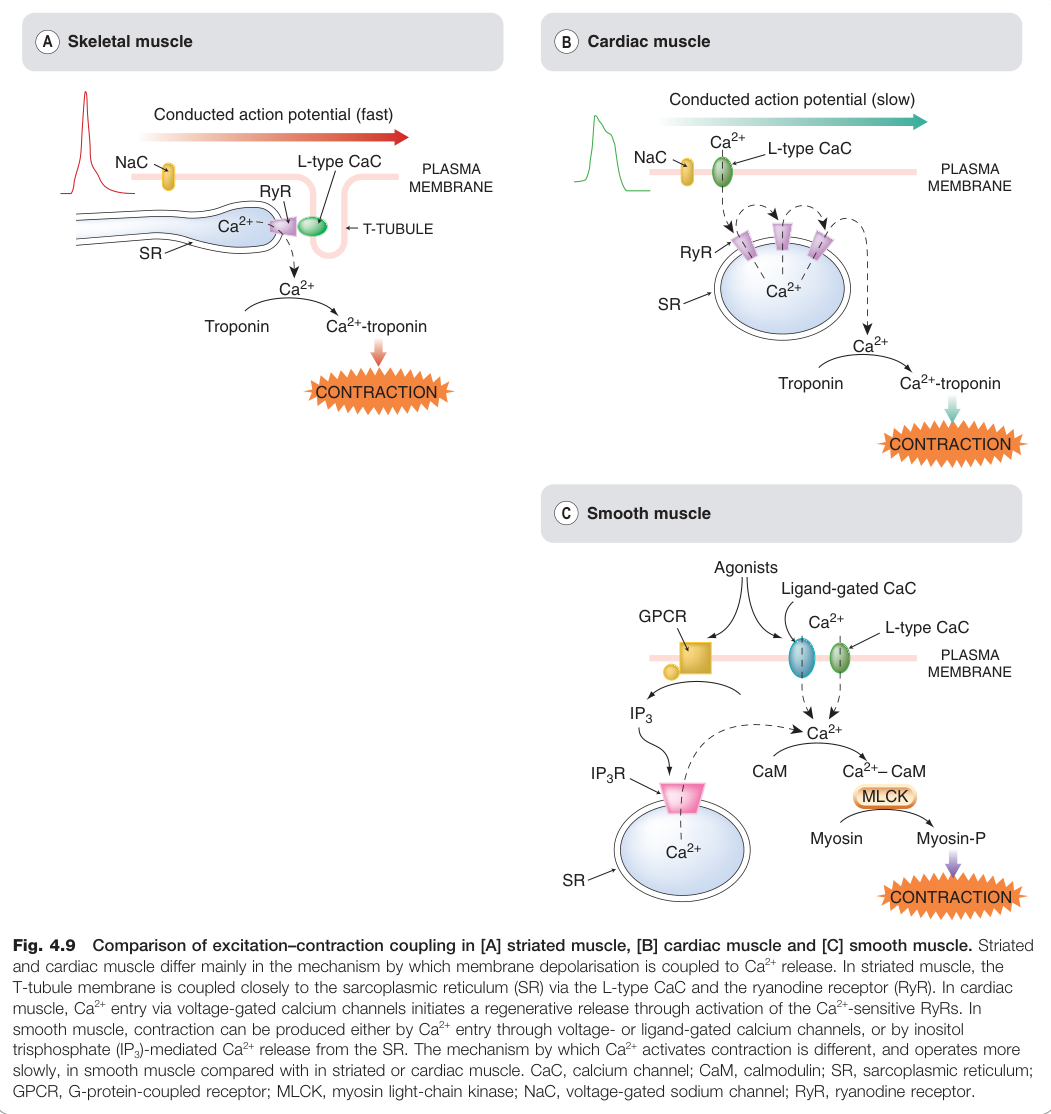
Figure 4.9:骨骼肌、心肌與平滑肌興奮–收縮偶聯機制的比較
心肌#
- 缺乏 T 管,漿膜與 SR 無直接偶聯
- 動作電位有特徵性「平台期」,期間 L 型 Ca²⁺ 通道持續開放,少量 Ca²⁺ 進入
- 進入的 Ca²⁺ 再激活 SR 上的 RyR(對 Ca²⁺ 敏感),引發全有全無的大量 Ca²⁺ 釋出(CICR)
- 最終 Ca²⁺ 同樣透過肌鈣蛋白啟動收縮
心肌收縮力高度依賴動作電位持續時間及 L 型 Ca²⁺ 通道的功能。地高辛透過升高 [Ca²⁺]i 增強心肌收縮力,機制為抑制 Na⁺–K⁺-ATPase → 升高 [Na⁺]i → 削弱 Na⁺–Ca²⁺ 交換子的排 Ca²⁺ 能力。
平滑肌#
平滑肌機制最為複雜,Ca²⁺ 訊號可來自多種途徑:
- Ca²⁺ 進入途徑:L 型 Ca²⁺ 通道(動作電位介導)、P2x 受體(ATP 配體閘控)
- Ca²⁺ 釋出途徑:GPCR → IP₃ → IP₃R → SR 釋放 Ca²⁺(不需動作電位)
收縮的分子機制與橫紋肌不同:
- Ca²⁺ → 鈣調蛋白 → 活化肌凝蛋白輕鏈激酶(myosin light-chain kinase, MLCK)
- MLCK 磷酸化肌凝蛋白輕鏈 → 收縮
- **肌凝蛋白磷酸酶(myosin phosphatase)**去磷酸化 → 鬆弛
- MLCK 與磷酸酶均受 cAMP 和 cGMP 調節
許多臨床重要藥物(心血管、呼吸道、消化道)透過促進或抑制平滑肌收縮發揮效果,目標包括 Ca²⁺ 通道、K⁺ 通道、GPCR 及環核苷酸路徑(見圖 4.10)。
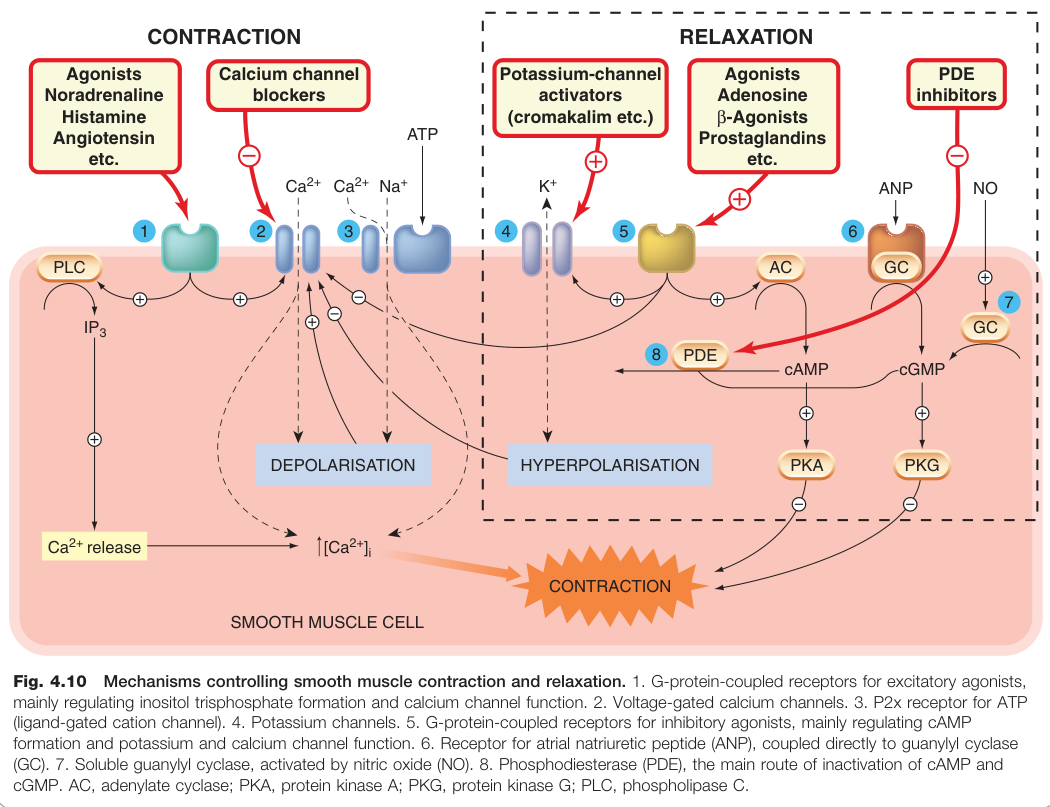
Figure 4.10:平滑肌收縮與鬆弛的調控機制,包括 GPCR、IP₃、Ca²⁺ 通道、MLCK 及環核苷酸路徑
化學傳遞物的釋放#
囊泡外胞作用(Exocytosis)#
大多數化學傳遞物(神經傳遞物、激素、細胞激素)儲存於囊泡(vesicle),透過**外胞作用(exocytosis)**釋出:
- [Ca²⁺]i 上升 → Ca²⁺ 結合囊泡蛋白突觸素(synaptotagmin)
- 促進突觸泡蛋白(synaptobrevin)與漿膜的突觸蛋白(syntaxin)結合,即 SNARE 複合體(SNARE complex)
- 囊泡膜與漿膜融合,釋出內容物(量子釋放)
- 空囊泡被內胞作用(endocytosis)回收、重新裝載傳遞物
快速傳遞物(如麩胺酸)的囊泡停靠在主動區(active zone)靠近 Ca²⁺ 通道處,逐衝動釋放;慢速傳遞物(如神經肽)的囊泡較遠,需高頻放電後累積足夠 Ca²⁺ 才釋放。
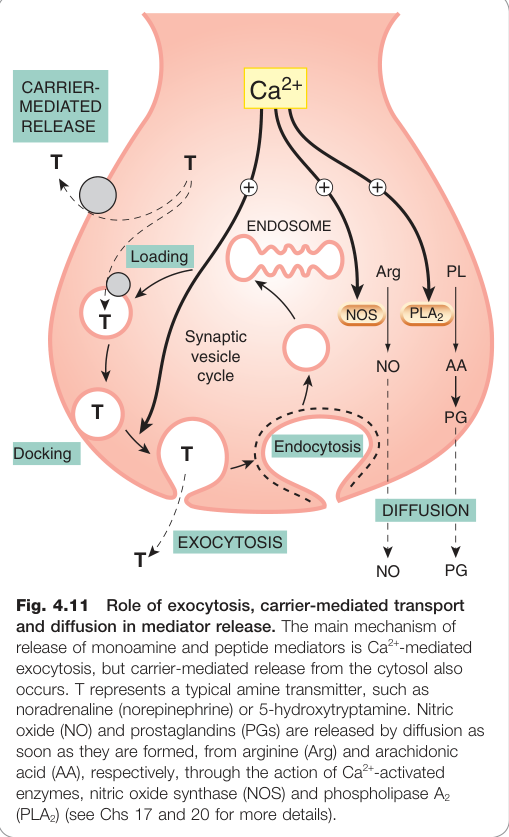
Figure 4.11:外胞作用、載體介導轉運與擴散在傳遞物釋放中的角色,以及單胺類和肽類傳遞物的主要釋放機制
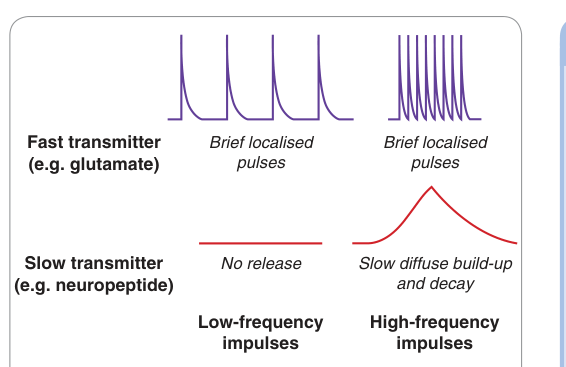
Figure 4.12:快速(如 glutamate)與慢速(如神經肽)傳遞物釋放的時程與頻率依賴性,反映囊泡停靠位置與 Ca²⁺ 通道距離的差異
非囊泡釋放機制#
- 載體介導釋放:安非他命(amphetamines)將胺類從囊泡置換至細胞質,再藉單胺轉運蛋白逆向轉運至胞外,不依賴 Ca²⁺
- 擴散性傳遞物:一氧化氮(NO)與前列腺素(prostaglandins)合成後直接擴散釋出,合成速率受 [Ca²⁺]i 調控
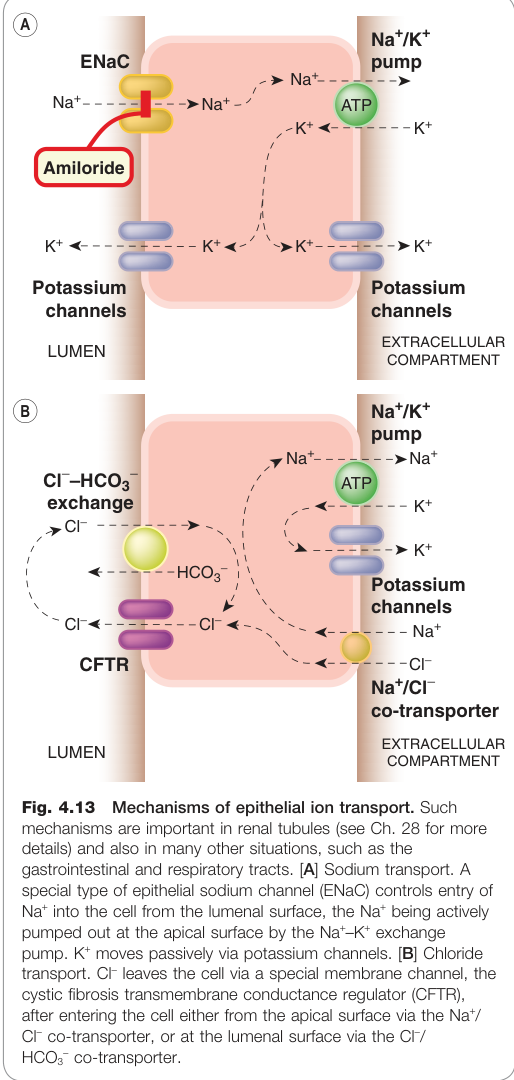
上皮離子轉運#
液體分泌上皮(腎小管、唾液腺、氣道、消化道)具有兩種主要離子轉運機制:
Na⁺ 轉運#
- **上皮型鈉離子通道(ENaC)**負責 Na⁺ 從腔內側進入細胞
- Na⁺–K⁺-ATPase 從細胞另一側主動輸出 Na⁺,水分被動跟隨
- **醛固酮(aldosterone)**透過核受體增加 ENaC 表達,促進腎臟 Na⁺ 重吸收
- **阿米洛利(amiloride)**選擇性阻斷 ENaC,用作利尿劑
Cl⁻ 轉運#
- 關鍵分子:囊性纖維化跨膜傳導調節蛋白(CFTR)——Cl⁻ 離子通道
- CFTR 突變導致囊性纖維化(cystic fibrosis),影響氣道分泌功能
- CFTR 受 cAMP 活化(透過蛋白激酶 A 磷酸化)
- 霍亂毒素等透過大量升高 cAMP → CFTR 過度活化 → 大量腸液分泌 → 腹瀉
Na⁺ 與 Cl⁻ 轉運均受 Ca²⁺ 與 cAMP 等第二訊息調節;Ca²⁺ 升高通常也能刺激分泌,可能也透過 CFTR 途徑作用。