章節概覽#
本章是周邊神經藥理學的奠基章節,闡述自主神經系統(autonomic nervous system, ANS)與軀體運動神經系統的解剖組織、神經傳導(neurotransmission)的化學機制,以及膽鹼性(cholinergic)與腎上腺素性(adrenergic)傳遞的完整分子細節。理解神經傳導的每一個步驟,是後續四章(蕈毒鹼受體、抗膽鹼酯酶、神經肌肉接合與神經節、腎上腺素性藥物)藥物作用機轉的共同基礎。
神經傳導的每一個步驟——合成、儲存、釋放、與受體結合、終止作用——都是潛在的藥物介入點(Table 8–7 系統性整理了各步驟的代表性藥物)。
自主神經系統的解剖組織#
ANS 調節不受意識控制的功能:呼吸、循環、消化、體溫、代謝、流汗與部分內分泌腺分泌,是維持內在環境恆定(homeostasis)的主要調控者。其傳出部分分為交感(sympathetic)與副交感(parasympathetic)兩大分支,多數內臟同時接受兩者支配。
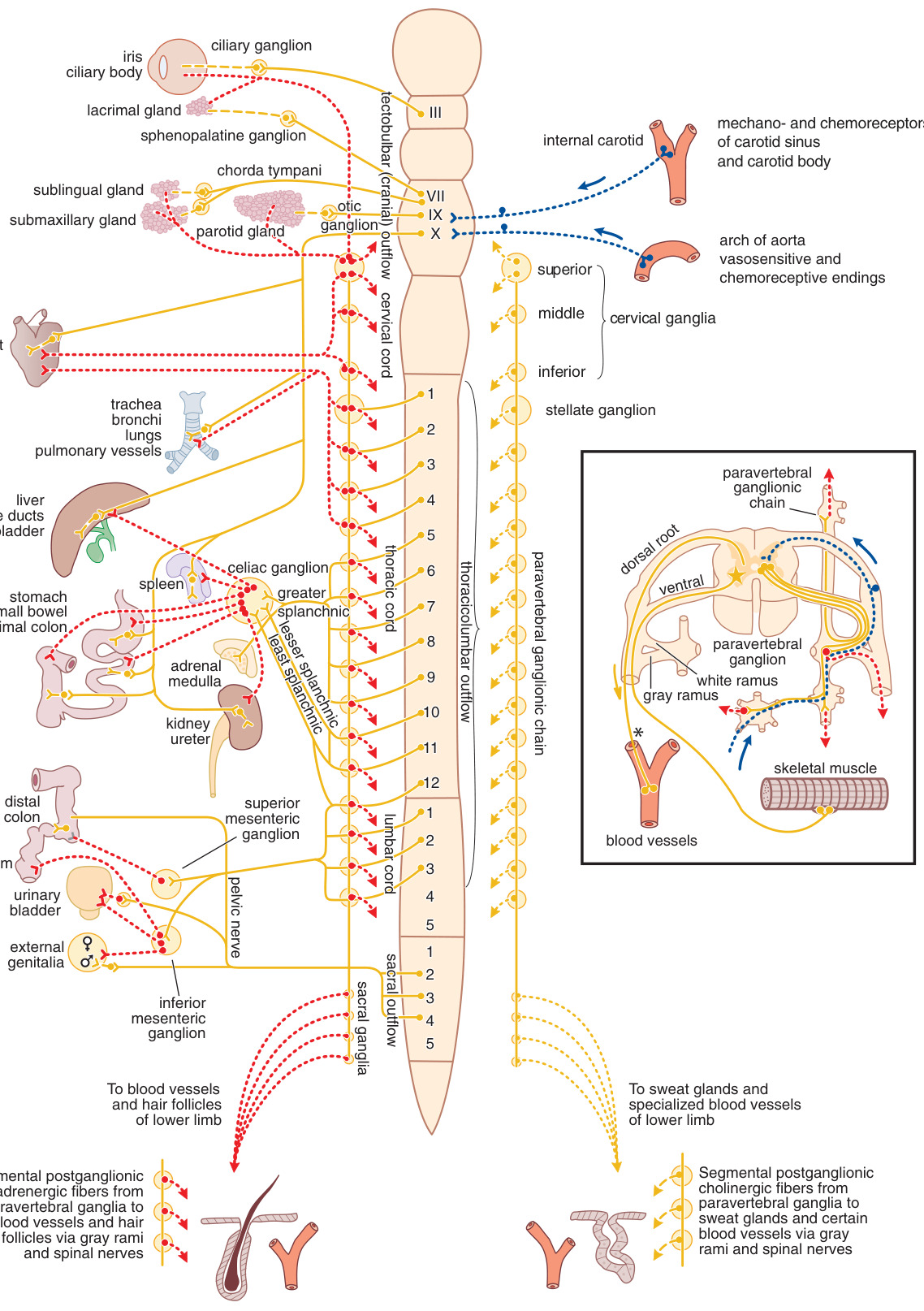
Figure 8–1:自主神經系統概觀
交感神經系統(胸腰段輸出)#
- 節前纖維(preganglionic fiber)源自胸髓到第三腰髓,經前根離開,與位於腦脊髓軸外的交感神經節(sympathetic ganglia)中的神經元形成突觸。
- 交感神經節分布於三處:椎旁(paravertebral,22 對形成兩側鏈)、椎前(prevertebral,如腹腔節 celiac、上下腸繫膜節)與終末(terminal)。
- 一條節前纖維可通過多個神經節並與大量節後神經元(postganglionic neuron)接觸,比例可達 1:20 以上,造就交感系統的瀰漫性放電(diffuse discharge)特性。
- 腎上腺髓質(adrenal medulla)在胚胎與解剖上類似交感神經節,由節前纖維(釋放乙醯膽鹼,acetylcholine)支配;其主要兒茶酚胺(catecholamine)為腎上腺素(epinephrine),約占釋放量的 80%。
副交感神經系統(顱薦段輸出)#
- 中樞起源於中腦、延腦與薦髓。
- 顱部輸出包含第三、七、九、十對腦神經的自主成分;迷走神經(vagus, 第十對)攜帶大量節前纖維至胸腹臟器壁內的小神經節。
- 副交感系統的節前纖維長、節後纖維短,終末神經節靠近或位於所支配的器官內,故影響較侷限、放電較離散。
腸神經系統(enteric nervous system, ENS)#
- 由肌間(Auerbach’s)神經叢與黏膜下(Meissner’s)神經叢組成,控制腸道的混合、推進與吸收。
- ACh 是興奮性運動神經元的主要傳遞物質(作用於 M2、M3 受體),另有神經激肽(neurokinins)參與興奮;抑制性運動神經元則經由一氧化氮(nitric oxide, NO)、ATP(作用於 P2Y1)與血管活性腸肽(VIP)等介導鬆弛。
- 運動神經元多透過 Cajal 間質細胞(interstitial cells of Cajal, ICCs)間接支配平滑肌。
交感與副交感的傳遞物質在多數器官互為「生理性拮抗」——一方抑制某功能、另一方則增強之。器官在任一時刻的活動,反映兩分支整合後的淨效果(詳見 Table 8–1 對各效應器官反應的完整對照)。
神經傳導的步驟#
神經衝動透過釋放特定化學傳遞物質,引發平滑肌、心肌、骨骼肌、外分泌腺與突觸後神經元的反應。
軸突傳導(axonal conduction)#
- 靜止電位約為內側 -70 mV,主要由膜對 K⁺ 的高通透性與細胞內外離子梯度(由 Na⁺,K⁺-ATPase 維持)決定。
- 去極化達閾值後啟動動作電位(action potential):先是電壓敏感性 Na⁺ 通道開啟造成快速去極化,繼而 Na⁺ 通道去活化與 K⁺ 通道延遲開啟造成再極化。
- 河豚毒素(tetrodotoxin)與蛤蚌毒素(saxitoxin)藉阻斷電壓敏感性 Na⁺ 通道而阻斷軸突傳導;箭毒蛙毒素(batrachotoxin)與蠍毒則造成持續去極化。
接合傳導(junctional transmission)#
- 傳遞物質的儲存與釋放:小分子傳遞物質儲存於突觸小泡。動作電位使軸突終末去極化,引發 Ca²⁺ 內流,促使小泡膜與軸漿膜融合並胞吐(exocytosis)。SNARE 蛋白複合體(synaptobrevin/VAMP、syntaxin、SNAP-25)介導融合;肉毒桿菌毒素(botulinum toxin)與破傷風毒素藉水解這些蛋白阻斷釋放。
- 與接合後受體結合並產生接合後電位:可形成興奮性突觸後電位(EPSP,陽離子通透性增加)或抑制性突觸後電位(IPSP,Cl⁻ 或 K⁺ 通透性增加導致超極化)。
- 啟動接合後活動:EPSP 超過閾值即引發傳導性動作電位或肌肉收縮。
- 傳遞物質的破壞或消散:膽鹼性突觸藉乙醯膽鹼酯酶(acetylcholinesterase, AChE)快速水解;正腎上腺素(norepinephrine, NE)主要藉再攝取與擴散終止作用。
- 非電生理功能:傳遞物質持續微量釋放可能參與神經滋養性(trophic)調控。
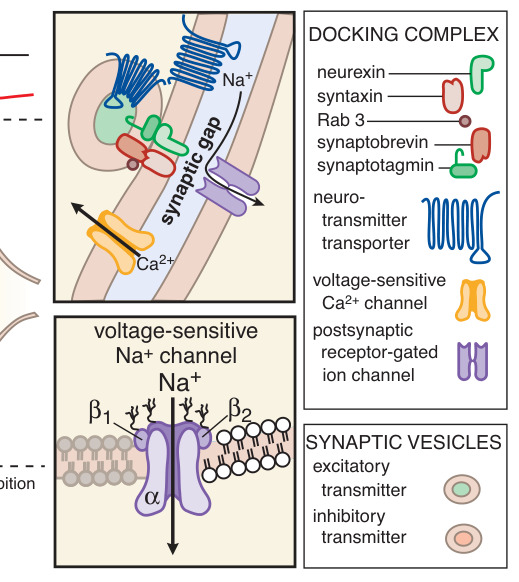
Figure 8–3:興奮性與抑制性神經傳遞的步驟
突觸前受體(presynaptic receptors)#
- 自體受體(autoreceptors):神經元自身傳遞物質回饋調控自身的合成與釋放(如 NE 經 α2A、α2C,ACh 經 M2、M4)。
- 異體受體(heteroreceptors):對鄰近神經元釋放的物質起反應(如 NE 可經 α2 抑制 ACh 釋放,ACh 可經 M2、M4 抑制 NE 釋放)。
膽鹼性傳遞(cholinergic transmission)#
ACh 是大多數節後副交感纖維、少數節後交感纖維、交感與副交感的節前纖維、骨骼肌運動神經,以及部分中樞神經元的傳遞物質。
合成、儲存與釋放#
- **膽鹼乙醯轉移酶(choline acetyltransferase, ChAT)**催化膽鹼(choline)與乙醯輔酶 A(acetyl CoA)合成 ACh。
- 膽鹼攝取是合成的限速步驟,由高親和力、Na⁺/Cl⁻ 依賴的轉運體(CHT1)負責,可被 hemicholinium-3 抑制。
- ACh 經囊泡乙醯膽鹼轉運體(VAChT)儲入小泡,此過程可被 vesamicol 抑制。
- 釋放藉 Ca²⁺ 依賴的胞吐進行,可被肉毒桿菌毒素阻斷。
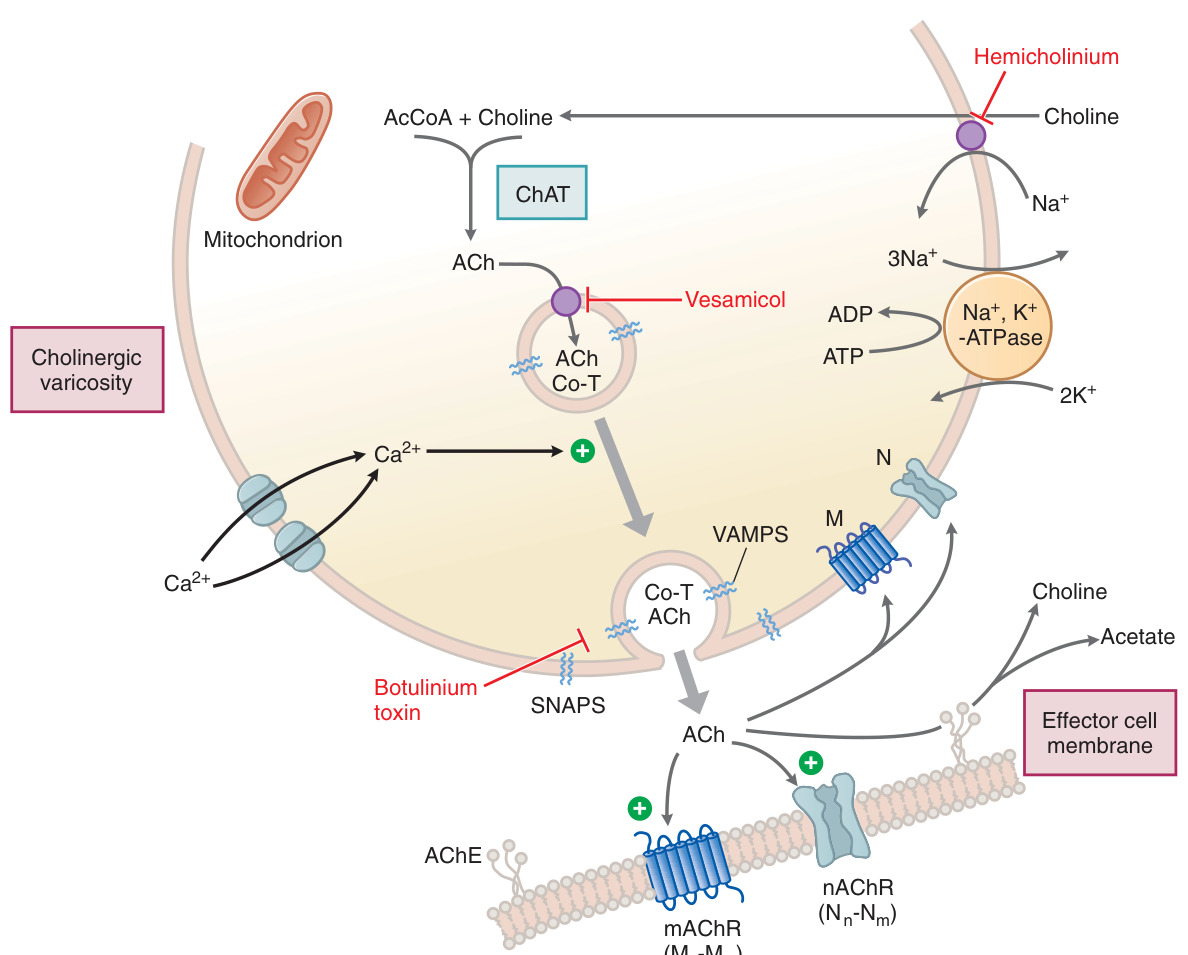
Figure 8–4:膽鹼性神經效應器接合處
乙醯膽鹼酯酶(AChE)#
- 高度集中於神經肌肉接合的突觸後終板,水解 ACh 為膽鹼與醋酸,速度極快(<1 ms)。
- 丁醯膽鹼酯酶(butyrylcholinesterase, BuChE,又稱 pseudocholinesterase)主要由肝臟合成、存在於血漿,水解攝入的酯類。
- 幾乎所有抗膽鹼酯酶藥物的效應都源自抑制 AChE 而使 ACh 累積(詳見抗膽鹼酯酶藥物一章)。
膽鹼性受體與訊號傳導#
Sir Henry Dale 區分了 ACh 的「菸鹼樣作用(nicotinic)」與「蕈毒鹼樣作用(muscarinic)」,此分類至今仍是膽鹼性受體的主要分野。
菸鹼性受體(nicotinic receptors, nAChRs)#
- 為配體閘控離子通道(ligand-gated ion channels),由五個同源次單元環繞中央孔組成;活化後迅速增加 Na⁺、Ca²⁺ 通透性,造成去極化與興奮。
- 分為肌肉型(骨骼肌神經肌肉接合,成人 (α1)₂β1εδ,胎兒 (α1)₂β1γδ)與神經元型(自主神經節、腎上腺髓質、CNS)。
- 神經元型 nAChRs 在 CNS 多扮演突觸調節角色,調控 ACh、DA、NE、glutamate、5-HT 等的釋放。
蕈毒鹼性受體(muscarinic receptors, mAChRs)#
- 為 G 蛋白偶聯受體(GPCRs),反應較慢,可興奮或抑制。哺乳類有五種亞型(M1–M5):
- M1、M3、M5:偶聯 Gq/11,活化磷脂酶 C(PLC),生成 IP3(釋放胞內 Ca²⁺)與 DAG(活化 PKC),造成平滑肌收縮與腺體分泌。
- M2、M4:偶聯 Gi/Go,抑制腺苷酸環化酶(adenylyl cyclase)、活化內向整流 K⁺ 通道、抑制電壓閘控 Ca²⁺ 通道,造成超極化與抑制(在心肌負責 ACh 的負性變時、變力效應)。
- 各亞型分布與疾病關聯(Table 8–3):M2 為心臟主要亞型;M3 為膀胱逼尿肌主要亞型;M1、M4、M5 富集於 CNS。
腎上腺素性傳遞(adrenergic transmission)#
涵蓋三種兒茶酚胺:NE(多數交感節後纖維的主要傳遞物質)、DA(錐體外系與中皮質、中邊緣路徑的主要傳遞物質)與 epinephrine(腎上腺髓質主要荷爾蒙)。
合成(Figure 8–5)#
- 酪胺酸(tyrosine)→ DOPA → 多巴胺(dopamine)→ NE →(僅在腎上腺髓質與少數腦幹神經元)epinephrine。
- **酪胺酸羥化酶(tyrosine hydroxylase, TH)**催化的羥化是限速步驟,受兒茶酚化合物回饋抑制。
- 多巴胺 β-羥化酶(DβH)將 DA 轉為 NE;苯乙醇胺-N-甲基轉移酶(PNMT,受糖皮質素誘導)將 NE 甲基化為 epinephrine。
儲存、釋放與再攝取#
- 兒茶酚胺由囊泡單胺轉運體(VMAT2)儲入小泡,VMAT2 可被 reserpine 抑制,導致兒茶酚胺耗竭。
- 釋放藉 Ca²⁺ 依賴胞吐進行,並與共傳遞物質(co-transmitters)神經肽 Y(NPY)與 ATP 一同釋放。
- 再攝取是終止 NE 作用的主要機制:NE 轉運體(NET,原稱 uptake 1)回收約 87% 釋出的 NE,可被 cocaine 與三環抗憂鬱劑(如 imipramine)選擇性阻斷。
- 間接作用型擬交感胺(如 tyramine、ephedrine、amphetamine)為 NET 受質,藉置換出儲存的 NE 而作用,並易產生快速耐受(tachyphylaxis)。
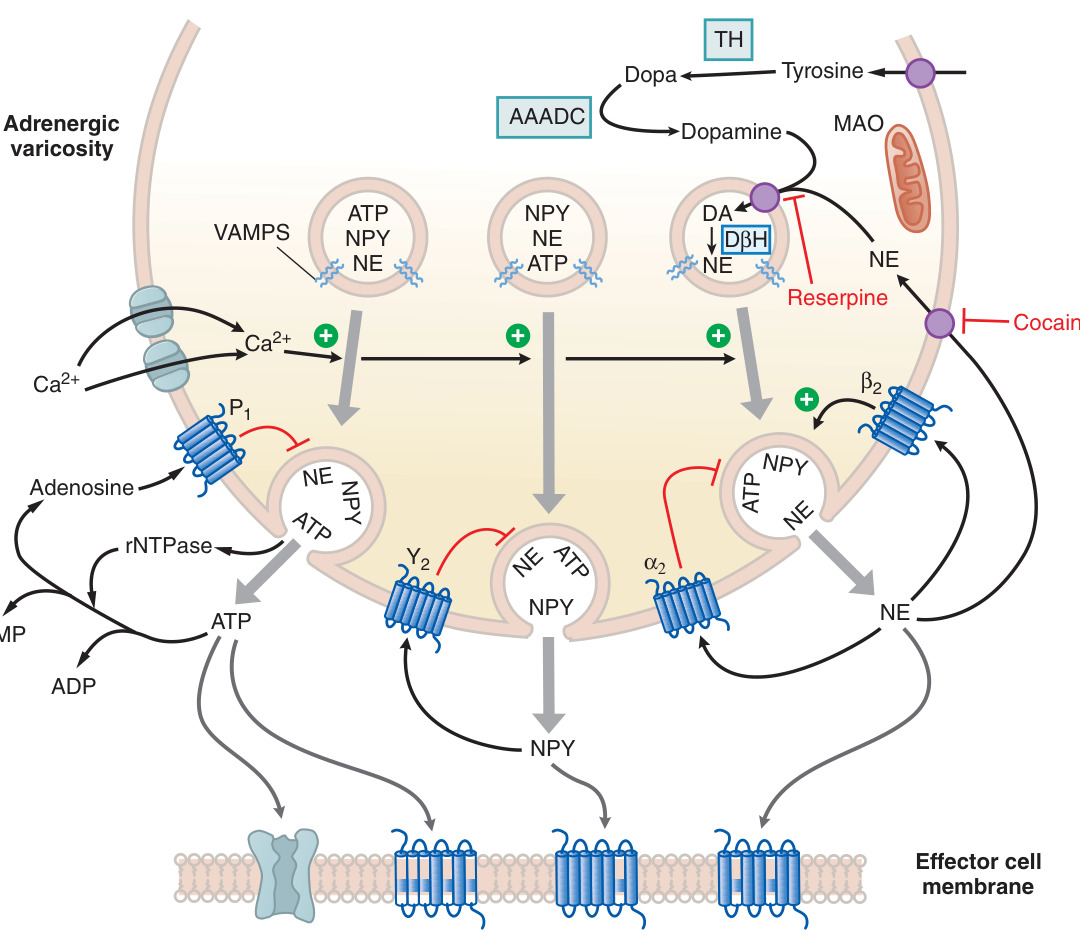
Figure 8–6:腎上腺素性神經效應器接合處
作用的終止與代謝(Figure 8–7)#
- 兩種關鍵代謝酶:單胺氧化酶(monoamine oxidase, MAO)(位於粒線體外膜,含 MAO-A、MAO-B 同功酶)與兒茶酚-O-甲基轉移酶(catechol-O-methyltransferase, COMT)(主要在肝、腎,多為胞質型)。
- NE 與 epinephrine 的最終代謝產物為香草扁桃酸(vanillylmandelic acid, VMA)。
不可逆 MAO-A 抑制劑(如 phenelzine、tranylcypromine)會增加食物中 tyramine 的生體可用率,誘發 NE 大量釋放而導致高血壓危象(hypertensive crisis)。選擇性 MAO-B 抑制劑(如 selegiline)或可逆性 MAO-A 抑制劑(如 moclobemide)較不易引發此交互作用。
腎上腺素性受體的分類#
Ahlquist 依據致效劑的效價順序首先提出 α 與 β 兩型受體。
- β 受體致效劑效價順序:isoproterenol > epinephrine ≥ norepinephrine;可被 propranolol 阻斷。再分為 β1(心肌,epinephrine 與 NE 等效)、β2(平滑肌等,epinephrine 強 10–50 倍)與 β3(脂肪組織等,產熱與脂解)。三者皆偶聯 Gs、活化腺苷酸環化酶、提升 cyclic AMP。
- α 受體致效劑效價順序:epinephrine ≥ norepinephrine » isoproterenol;可被 phenoxybenzamine 阻斷。
- α1(α1A、α1B、α1D):偶聯 Gq,活化 PLC,介導血管平滑肌收縮(α1A 為多數血管床主要亞型)。
- α2(α2A、α2B、α2C):偶聯 Gi,抑制腺苷酸環化酶、降低 cAMP。α2A 為交感神經末梢主要的抑制性自體受體,CNS 中 α2A 活化降低交感外流(clonidine 降血壓的機轉之一)。
反覆暴露於腎上腺素性致效劑會導致反應逐漸減弱,稱為去敏感化(desensitization)或快速耐受(tachyphylaxis)。機制包括 GRK 與 PKA/PKC 對受體的磷酸化、β-arrestin 結合與訊號解偶聯、受體內吞與下調等;β2 受體最易被調控。
神經與內分泌系統的關係#
神經 humoral 傳遞理論顯示神經系統與內分泌系統的深層相似性。ANS 負責對環境變化的快速調整(作用短暫、侷限於釋放處);內分泌系統則藉荷爾蒙進入循環,在遠端廣泛部位作用較慢、較全面的適應。兩者皆在下丘腦整合。
藥理介入分類#
各神經傳導步驟對應的藥物(Table 8–7):
- 干擾傳遞物質合成:hemicholinium(膽鹼性)、α-methyltyrosine/metyrosine(抑制 TH,腎上腺素性)。
- 以相同路徑代謝為偽傳遞物質:methyldopa →α-methyl-NE(α2 致效劑)。
- 阻斷神經末梢膜轉運:cocaine、imipramine(使 NE 在受體處累積)。
- 阻斷儲存小泡轉運:vesamicol(ACh)、reserpine(NE,導致耗竭)。
- 促進釋放或置換:latrotoxins(ACh)、amphetamine/tyramine(擬交感)。
- 阻止釋放:botulinum toxin(ACh,臨床用於肌張力異常、斜視、眼瞼痙攣與美容除皺)、bretylium/guanadrel(腎上腺素性)。
- 模擬傳遞物質(致效劑)與阻斷接合後受體(拮抗劑):涵蓋蕈毒鹼、菸鹼、α 與 β 各亞型的選擇性藥物。
- 抑制酶性分解:AChE 抑制劑(如 neostigmine、pyridostigmine、edrophonium);MAO 與 COMT 抑制劑。