核心主題#
哺乳動物的存活仰賴體內恆定(homeostasis):氧、二氧化碳、水、鹽、能量、體溫等都需維持在狹窄範圍。本章探討身體如何透過神經與行為機制維持恆定,並聚焦於能量平衡作為理解所有調節系統的範例。
「能量即愉悅(Energy is delight)」並非詩意誇張——感覺愉悅的神經機制,是腦把「我們正接觸到對生物有用的物事」這個訊息告訴自己的方式。
與恆定相關的感受狀態#
加爾各答黑洞事件(1756)的軍官回憶錄被引用為極端飢渴經驗的縮影:
- 強烈口渴將心智「漏斗化」為單一目標:水
- 倖存者甚至吸吮自己襯衫上的汗水以舒緩
- 這展現了內在身體失衡如何徹底接管腦中的目標與行動
強烈感受不只主觀體驗,更是腦的價值編碼裝置:以「飢、渴、痛、愉」標記哪些事件對生存重要。
不被意識到的快速調節:呼吸與「窒息警報」#
- 呼吸由腦幹自動調節,平時不需意識參與
- 腦並不直接偵測缺氧,而是監測 CO₂ 累積與血液酸鹼變化
- 一旦呼吸節律被中斷(窒息、勒頸),即觸發窒息警報反射(suffocation alarm reflex)
- 這個原始迴路可能就是人類**恐慌症發作(panic attacks)**的部分根源
共用一條 SEEKING 系統,搭配多個專屬偵測器#
身體偵測器(醫學/物理量) 共用驅動系統 消費反射
───────────────────── ───────────── ──────────────
glucose、osmolality、體溫、 ────→ SEEKING ────→ 舔、咬、嚼、吞
性荷爾蒙、CO₂、體積…… (LH 主軸) (具體針對需求)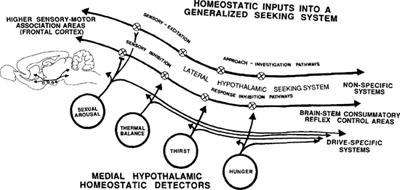
Figure 9.1: How medial hypothalamic regulatory detectors access a shared SEEKING system
- 演化不可能為每種需求建立一條獨立的「追尋系統」
- **內側下視丘(medial hypothalamus)**有大量化學敏感神經元(葡萄糖、脂肪酸、滲透壓、體溫、性類固醇等的偵測器)
- 它們透過軸突連到**腹側被蓋區(VTA)**與外側下視丘,調控 SEEKING 系統的活躍度
- 不同需求只需「動」同一個尋找腳本,加上特化的消費反射
老鼠的「LH 自我刺激區」在貓身上往往對應獵食性攻擊。同一系統的外顯行為視物種而異,這也是早期文獻把 LH 系統誤判為「獎賞中樞」的另一例。
「驅力(drive)」概念的式微#
- 傳統「飢渴是抽象內在驅力」的說法已被認為過於模糊
- 取而代之的是身體需求狀態(need state)+ 誘因(incentive)+ SEEKING 系統的三角結構
- 神經迴路本身對誘因的反應比外在物的物理屬性更直接
進食的真實調節:是能量,不是個別餐點#
實驗(圖 9.2):
- 把每天 25% 卡路里以營養液直接注入胃,無論是脂肪、蛋白還是碳水
- 動物會自動把口攝食減少 25%
- 結論:主要被調節的是「能量含量」——管它從什麼大分子來
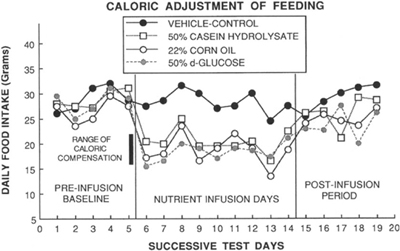
Figure 9.2: Daily food intake when 25% of calories are infused directly into the stomach
食物熱量密度提升 → 攝食量下降
熱量密度稀釋 → 攝食量上升
胃內補充熱量 → 口攝量等比例下降- 飽足是長期、跨越多餐的調節,單一餐次資訊有限
- 「超市飲食」(持續供應多樣可口食物)會讓大鼠和人類都過食肥胖
- 規律運動則有助維持體重穩定
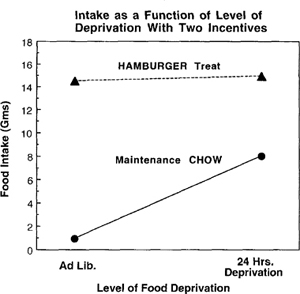
Figure 9.3: High-incentive (raw hamburger) vs. low-incentive food intake by deprivation level
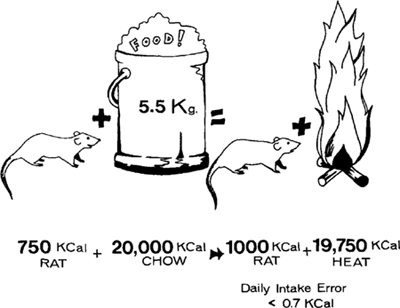
Figure 9.4: A female rat's approximate yearly energy balance equation
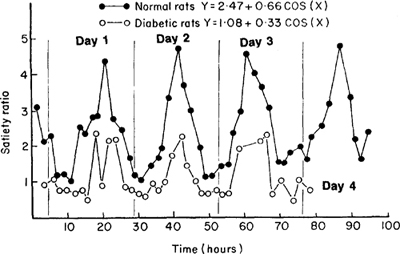
Figure 9.5: Average daily feeding cycle in normal vs. diabetic rats
鑑別「真正的飽足」與「假性食欲降低」#
許多藥物會抑制食欲,但多數可能只是讓動物不舒服而非引發真正飽足:
- 主要鑑別工具:條件性味覺嫌惡(conditioned taste aversion, CTA)——若藥物使動物隨後拒絕某新食物,多半是讓牠覺得「噁心」
- 例:CCK(cholecystokinin)一度被視為飽足訊號,但仔細測試後發現其實透過迷走神經傳遞胃腸道不適
- 「飽足後行為」(理毛 → 探索 → 小睡)這套舊指標太鬆散
- 潘克塞普建議使用幼鼠遊戲行為作為飽足的正向指標:
- 飢餓會大幅壓抑遊戲
- 真正的飽足應該「讓玩耍回來」
- CCK、GLP-1、opiate 拮抗劑等多數「飽足肽」皆無法讓遊戲恢復——意味它們其實只是不適或焦慮的誘因
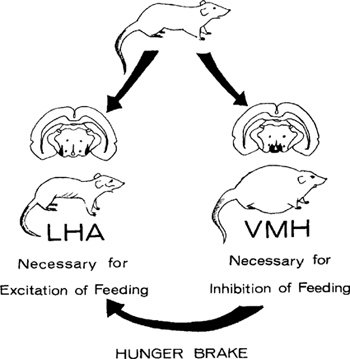
Figure 9.7: Regulatory effects of lateral (left) and medial (right) hypothalamic damage
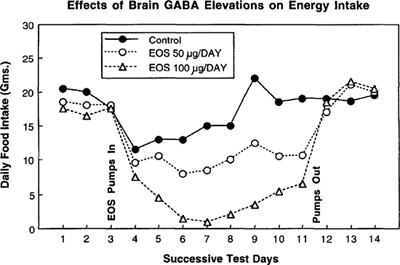
Figure 9.9: Daily food intake during GABA breakdown inhibition
長期能量恆定與瘦素#
- 真正參與長期能量調節的關鍵在脂肪細胞分泌的瘦素(leptin)
- 瘦素將「身體儲能總量」的訊息送到下視丘
- 大多數現有減肥藥只作用於短期食欲機制,因此效果有限
- 真正能用的長期方案,可能要操弄瘦素受體或其下游訊號
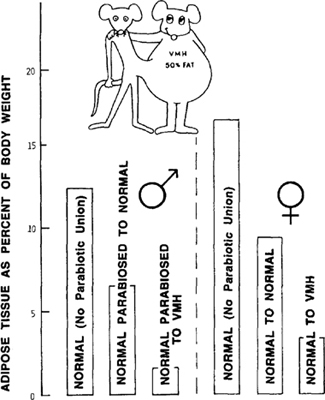
Figure 9.6: Body fat changes in parabiotic union of normal rats
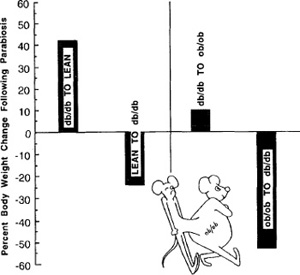
Figure 9.8: Body weight changes in genetically obese (db/db, ob/ob) parabiotic mice
感覺愉悅:身體恆定的訊號讀取#
- 飢餓時甜食特別好吃;飽足後同樣甜食的愉悅感大幅降低——這是「alliesthesia」現象
- 同理適用於性、觸覺、溫度
- 新生兒對四種基本味道(甜、酸、苦、鹹)的反射性面部表情非常明顯
- 大鼠對甜溶液舔脣、對苦溶液搖頭並抹下巴——可作為皮質下情感反應的客觀指標
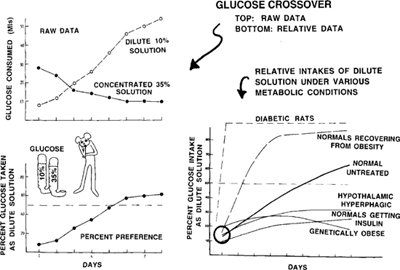
Figure 9.10: Sugar water consumption patterns with two solution concentrations
各種「感覺愉悅」很可能不來自不同的快感系統,而是來自**同一群神經化學(特別是腦內鴉片肽 endogenous opioids)**作用於略有不同的腦區,加上各自的感覺特徵。
快感的神經化學:腦內鴉片肽是主角#
- 給予非熱量甜味劑(saccharin)時,動物的攝食隨時間下降;若加上鴉片受體拮抗劑 naltrexone,下降更快
- 甜的物質會促進腦內鴉片肽釋放——這也是嬰兒在喝糖水後減少哭叫、痛閾上升的原因
- 與味道、配對的腦內鴉片活化/阻斷可創造條件性味道偏好
- 性、母性、社會接觸的愉悅也都有鴉片成分
- 高劑量鴉片大幅降低食欲與其他多種獎賞動機;低劑量則微微激活 SEEKING——進食上升
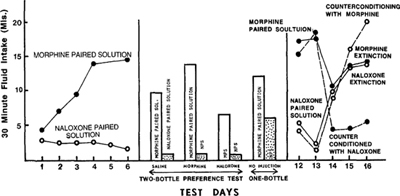
Figure 9.11: Saccharin solution intake — morphine/naloxone conditioned preference
其他調節系統#
- 飲水:兩種口渴
- 滲透壓口渴:偵測細胞內鈉濃度,由 OVLT(lamina terminalis 的血管器)感應
- 容量性口渴:偵測血流容量下降,主要由穹窿下器官(subfornical organ)感應;與肺—腎所釋的 angiotensin 相連
- 鈉:因鈉是稀缺生命元素,腦會記憶「鹽存在的地點」,即便當下不缺鈉;缺鈉時這份記憶會立刻被喚醒
- 微量營養素/維生素:難以味覺辨識,主要透過進食後感覺學習
- 生病感:免疫細胞激素(cytokines,例:interleukins)作用於腦的特定區域(後極區 area postrema、顳葉內側等)
Afterthought 重點#
- 短期食欲控制機制與長期能量平衡機制必須分清,否則藥物開發會走錯方向
- 飽足是一個內在訊號:身體能量已被恢復,並回饋給 SEEKING 系統使其降溫
- 真正有效的減重藥物,可能必須模擬「長期已飽」這個神經訊號,而非短期抑制進食